Медиальный подошвенный нерв, n. plantaris mеdialis, больше, чем латеральный.
НЕРВНАЯ СИСТЕМА
Нервная система человека (рис. 293) подразделяется на центральную (головной и спинной мозг) ипериферическую(нервные корешки, узлы, сплетения, черепные и спинномозговые нервы).
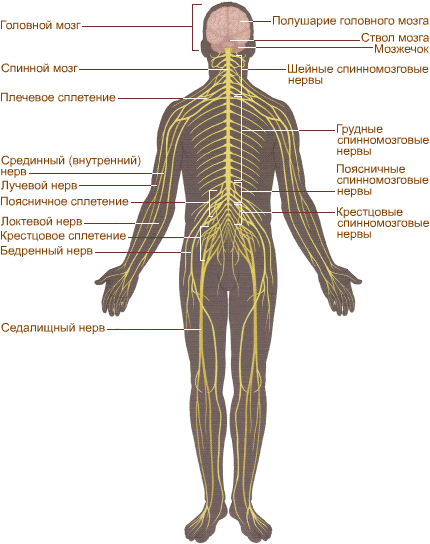
Рисунок 293. Схема нервной системы человека
На рисунке схематично показана нервная система человека. Она играет главную роль в согласованной деятельности организма. Особый комплекс периферических нервов и нервных узлов, иннервирующих сердце, легкие, пищеварительный тракт и другие внутренние органы, сосуды и ткани, и есть вегетативная нервная система. Ее работа, как правило, не зависит от волевых усилий человека, и в норме мы не ощущаем раздражения вегетативных узлов и нервов. Нервные узлы симпатического отдела вегетативной нервной системы образуют симпатические нервные стволы, расположенные около спинного мозга, а нервные узлы другого отдела — парасимпатического – лежат во внутренних органах или около них.
Для слаженной деятельности различных частей такой сложной системы, как организм человека, необходимо координирующее устройство соответственной сложности. И в самом деле, нервная система, интегрирующая деятельность всех частей тела, является, несомненно, самой сложной из всех систем органов. Мышцы и железы животного или человека носят общее название эффекторов; глаза, уши и другие органы чувств называются рецепторами. Нервная система, состоящая из головного мозга, спинного мозга и проводящих путей, соединяет рецепторы с эффекторами и передает импульсы, или «сообщения», от первых ко вторым. Она способна делать это таким образом, что при раздражении того или иного рецептора должным образом реагирует надлежащий эффектор. Основными функциями нервной системы являются проведение импульсов и интеграция деятельности различных систем организма. Координирующие функции нервной системы, эндокринная регуляция и собственные регуляторные механизмы внутриклеточных ферментных систем (торможение и стимуляция активности ферментов, индукция и репрессия их синтеза) — все это факторы, способствующие гомеостазу, т.е. поддержанию постоянства внутренней среды организма.
Центральная нервная система (ЦНС) — это совокупность нервных образований спинного и головного мозга, обеспечивающих восприятие, обработку, передачу, хранение и воспроизведе-ние информации с целью адекватного взаимодействия организма и изменений окружающей среды, координации оптимальной работы органов, их систем и организма в целом.
Каждая из этих структур имеет морфологическую и функциональную специфику. Но, наряду с этим, у всех структур нервной системы есть ряд общих свойств и функций, к которым относятся: нейронное строение, электрическая и химическая синаптическая связь между нейронами, образование локальных сетей из нейронов, реализующих специфическую функцию, множественность прямых и обратных связей между структурами, способность нейронов всех структур к восприятию, обработке, передаче и хранению информации, преобладание числа входов для ввода информации над числом выходов, способность к параллельной обработке информации, способность к саморегуляции, функционирование на основе рефлекторного доминантного принципа.
Головной мозг является важнейшим отделом ЦНС, в нем различают стволовую часть и конечный мозг, включающего подкорковые или базальные ганглии и большие полушария.
Основные части головного мозга выделяются уже к 3-му месяцу эмбрионального развития, а к 5-му месяцу эмбриогенеза уже хорошо заметны основные борозды больших полушарий.
К моменту рождения общая масса головного мозга составляет около 400 г., причем у девочек он несколько меньше (388 и 391 у девочек и мальчиков соответственно). По отношению к массе тела мозг у новорожденного значительно больше, чем у взрослого. Так, если у новорожденного он составляет 1/8 массы тела, то у взрослого — 1/40. Наиболее интенсивно головной мозг человека развивается в первые два года постнатального развития. Затем темпы его роста снижаются, но продолжают оставаться высокими до 6-7 лет, к этому моменту масса мозга достигает уже 4/5 массы взрослого мозга. Окончательное созревание головного мозга заканчивается только к 17-20 годам. К этому возрасту масса мозга увеличивается по сравнению с новорожденными в 4-5 раз и составляет в среднем у мужчин 1400 г, а у женщин — 1260 г. Следует отметить, что абсолютная масса мозга не определяет непосредственно умственные способности человека.
Изменения размеров, формы и массы мозга сопровождается изменением его внутренней структуры. Усложняется строение нейронов, форма межнейронных связей, становится четко разграниченным белое и серое вещество, формируются различные проводящие пути головного мозга.
Развития мозга, как и других систем, идет гетерохронно. Раньше других созревают те структуры, от которых зависит нормальная жизнедеятельность организма на данном возраст-ном этапе. Функциональной полноценности достигают вначале стволовые, подкорковые и корковые структуры, регулирующие вегетативные функции организма. Эти отделы по своему развитию приближаются к мозгу взрослого человека уже к 2-4 годам постнатального периода.
Продолговатый мозг, мост, средний мозг, промежуточный мозг и мозжечок относятся к стволу мозга. В филогенетическом отношении это наиболее древние нервные структуры и поэтому их функции тесно связаны с регуляцией примитивных функциональных процессов.
В процессе онтогенеза созревание структур стволовой части головного мозга наиболее интенсивно происходит в первые два года жизни. Окончательное формирование этих структур, особенно промежуточного мозга, завершается только в 13-16 лет, когда заканчивается половое развитие подростков. Многие особенности низшей и высшей нервной деятельности у детей подросткового возраста объясняются функциональными свойствами промежуточного мозга и некоторых других подкорковых структур.
Наиболее молодым в филогенетическом отношении является конечный мозг. В его состав входят большие полушария и расположенные под ними скопления серого вещества в виде подкорковых или базальных ганглиев.
Большие полушария осуществляют регуляцию высших нервных функций, лежащих в основе всех психических процессов человека. Правое и левое полушарие тесно связаны между собой с помощью огромного количества нервных волокон, образующих мозолистое тело. Многие нервные процессы, выходящие из какой-либо точки одного полушария, проецируются в сим-метричную точку другого полушария. Таким образом, в нервной деятельности полушарий конечного мозга проявляется свойство билатеральной симметрии.
Существует предположение, что в процессе онтогенеза развитие парной деятельности полушарий идет от неустойчивой симметрии к неустойчивой асимметрии, и наконец, к устойчивой функциональной асимметрии. Это подтверждается развитием бимануальных действий человека, то есть особенностей двигательной деятельности левой и правой рук. Пока-зано, что к праворукости дети переходят с 2-4 лет. В этом возрасте правши составляют 38%, а к 4-6 годам — 75%. Иначе говоря, в значительной степени моторная асимметрия зависит от условий воспитания ребенка, но вместе с тем существует и наследственная предраспо-ложенность.
Темпы созревания левого и правого полушарий имеют половые особенности. Левое полу-шарие у девочек развивается быстрее, что свидетельствует о более раннем созревании доминантного полушария. Данный факт косвенно подтверждается также более быстрым развитием у девочек речи и некоторых показателей психомоторики.
Высшим центром регуляции и управления всей деятельностью организма, начиная от самых примитивных физиологических отправлений и кончая сложнейшими психическими процес-сами у человека, является кора головного мозга. Активное формирование полушарий мозга начинается с 12-й недели эмбриогенеза и интенсивно продолжается в первые годы постнаталь-ного развития, особенно до 2 лет. Клеточное строение, форма и расположение борозд и извилин приближается к взрослому мозгу в 7 лет. А в лобных долях это различие сглаживается только к 12 годам. Существует прямая зависимость между морфофункциональным созреванием лобных долей больших полушарий и формированием психических функций у детей. Окончательное созревание больших полушарий и коры мозга завершается к 20-22 годам.
Морфологический анализ процессов созревания КГМ ребенка на клеточном уровне свидетельствует о постоянном увеличении размеров высших первичных, вторичных и третич-ных зон КГМ в процессе постэмбрионального развития: чем больше возраст ребенка, тем большие размеры занимают эти корковые зоны, и тем сложнее становится психическая деятельность.
Таким образом, в процессе постнатального развития происходит совершенствование морфо-логического строения КГМ, а параллельно этому и совершенствование высшей нервной деятельности ребенка и его психических процессов. Например, поля двигательного центра речи достигают функциональной полноценности только к 7 годам, к этому возрасту они увеличи-ваются на 64-73% в сравнении с мозгом новорожденного. То же можно сказать и о корковых зонах, ответственных за интеграцию слуховых и зрительных раздражителей, что имеет большое значение в формировании речи.
Важные данные о функциональной зрелости коры и подкорковых образований мозга и участия их в восприятии афферентных сигналов в разные возрастные периоды получены при использовании электрофизиологических методов. Анализ имеющихся в литературе данных о характере фоновой и вызванной электрической активности мозга человека на разных этапах онтогенеза рассматривается в связи с проблемой созревания высших отделов центральной нервной системы.
В раннем постнатальном периоде наиболее функционально зрелыми являются мезодиэнце-фалические структуры мозга, определяющие ЭЭГ-картину глубокого сна и реакцию возбуждения у новорожденного ребенка. Ряд факторов свидетельствует, что кора больших полушарий начинает функционировать уже с момента рождения ребенка. Нервные элементы коры больших полушарий новорожденного способны продуцировать кратковременную ритмическую электрическую активность. Это выражается в виде: 1) наличия групп синхронизи-рованных ЭЭГ-колебаний в затылочных областях мозга в переходном от бодрствования ко сну состоянии; 2) реакции усвоения ритма световых мельканий, наблюдаемой с первых часов жизни ребенка; 3) наличия ритмического сенсорного разряда, регистрируемого в затылочной области коры при значительной стимуляции. Отмечается, что кора больших полушарий новорожденных вовлекается в реакцию при афферентных воздействиях. В этом случае наблюдаются как генерализованные изменения электрической активности, обусловленные возбуждением подкорковых неспецифических структур мозга, так и локальные вызванные ответы, свидетельствующие о поступлении сигнала в кору больших полушарий по специфическому афферентному пути. Наличие ответных специфических и неспецифических реакций на афферентное раздражение означает функционирование восходящих ретикулярных и таламокортикальных связей, посылающих сенсорную информацию в кору больших полушарий. Для оценки способности коры воспринимать приходящую информацию наибольший интерес представляет наличие с момента рождения ребенка начальной позитивности вызванного специфического ответа, свидетельствующее о непосредственном участии нейронов 3-го и 4-го слоев коры в приеме афферентного сигнала.
Подчеркивая функционирование коры больших полушарий в период новорожденности, следует иметь в виду и отличия в деятельности ее нервных элементов по сравнению со взрослыми. Одним из таких проявлений служит отсутствие синхронизированной ритмической активности во время бодрствования новорожденных. Устойчивая ритмика в ЭЭГ бодрствую-щих детей регистрируется только с 2-3 месяцев постнатальной жизни. Появление организован-ной ритмики в состоянии спокойного бодрствования отражает важный этап в созревании коры мозга ребенка. В этом возрасте исчезают архаические рефлексы, развивается оптомоторная пространственная координация.
В течение первого года жизни формируется строго ритмическая электрическая активность частотой 5 Гц с фокусом в затылочной области коры, которая может рассматриваться как аналог альфа-ритма взрослого человека. С возрастом отмечается прогрессивное учащение альфа-ритма, появление и стабилизация его в центральных областях коры. Формирование основного ритма электрической активности, отражающее морфофункциональное созревание нейронного аппарата коры больших полушарий, заканчивается к 16-18-летнему возрасту. Созревание нервных элементов коры больших полушарий проявляется также в эволюции специфически вызванных потенциалов. В процессе индивидуального развития ребенка отмечается укорочение временных параметров ответа, усложнение его компонентного состава и появление вызванных потенциалов в ассоциативных областях коры. Включения ассоциа-тивных зон в прием и переработку качественно специфической информации, вероятно, опре-деляет возможность синтеза интегрального образа раздражителей разного информационного значения.
Параллельно с изменениями ЭЭГ, обусловленными функциональным созреванием коры больших полушарий, отмечается уменьшение выраженности подкорковых знаков в ЭЭГ (тета - волн, билатеральных пароксизмальных разрядов, усиленных неспецифических ответов). Ослаб-ление подкорковых знаков в ЭЭГ с возрастом можно объяснить усилением тормозных влияний созревающей коры на подкорковые структуры.
Таким образом, выявляются определенные этапы функционального созревания коры и подкорковых структур мозга, специфика их взаимных влияний в различные возрастные периоды.
Нервная ткань состоит из двух разновидностей клеток: нервных (нейронов) и глиальных. Глиальные клетки вплотную прилегают к нейрону, выполняя опорную, питательную, секретор-ную и защитную функции.
Вся нервная система построена на нервной ткани. Нервная ткань состоит из нервных клеток (нейронов) и связанных с ними анатомически и функционально вспомогательных клеток нейроглии. Нейроны выполняют специфические функции, являясь структурно-функциональной единицей нервной системы. Нейроглия обеспечивает существование и специфические функции нейронов, выполняет опорную, трофическую (питательную), разграничительную и защитную функции.
Нейрон (нейроцит) получает, перерабатывает, проводит и передает информацию, закоди-рованную в виде электрических или химических сигналов (нервных импульсов).
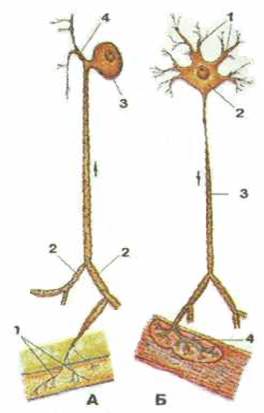
Рисунок 294. Нервные клетки. А — чувствительный нейрон. Б — двигательный нейрон. Стрелки показывают направление следования нервных импульсов. А: 1 - чувствительные нервные окончания; 2 - дендриты; 3 - тело нервной клетки; 4 - аксон. Б: 1 - дендриты; 1 - тело нервной клетки; 3 - аксон; 4 - двигательное нервное окончание (нервно-мышечная бляшка).
Каждый нейрон имеет тело, отростки и их окончания (рис. 294). Снаружи нервная клетка окружена оболочкой (цитолеммой), способной проводить возбуждение, а также обеспечивать обмен веществ между клеткой и окружающей их средой. Тело нервной клетки содержит ядро и окружающую его цитоплазму (перикарион). Цитоплазма нейронов богата органеллами (субклеточными образованиями, выполняющими ту или иную функцию). Диаметр тел нейро-нов варьирует от 4-5 до 135 мкм. Форма тел нервных клеток тоже различная — от округлой, овоидной до пирамидальной. От тела нервной клетки отходят различной длины тонкие отростки двух типов. Один или несколько древовидно ветвящихся отростков, по которым нервный импульс приносится к телу нейрона, называют дендритом. У большинства клеток их длина составляет около 0,2 мкм. Единственный, обычно длинный отросток, по которому нервный импульс направляется от тела нервной клетки — это аксон, или нейрит.
По количеству отростковнейроны подразделяются на униполярные, би- и мульти-полярные клетки. Униполярные (одноотростчатые) нейроны имеют лишь один отросток. У человека такие нейроны встречаются лишь на ранних стадиях внутриутробного развития. Биполярные (двухотростчатые) нейроны имеют один аксон и один дендрит. Их разно-видностью являются псевдоуниполярные (ложноуниполярные) нейроны. Аксон и дендрит этих клеток начинаются от общего выроста тела и в последущем Т-образно делятся. Мультиполяр-ные (многоотросчатые) нейроны имеют один аксон и много дендритов, они составляют большинство в нервной системе человека. Нервные клетки динамически поляризованы, т.е. способны проводить нервный импульс только в одном направлении — от дендритов к аксону.
В зависимости от функции нервные клетки подразделяют на чувствительные, вставочные и эффекторные.
Чувствительные (рецепторные, афферентные) нейроны. Эти нейроны своими окончаниями воспринимают различные виды раздражений. Возникшие в нервных окончаниях (рецепторах) импульсы по дендритам проводятся к телу нейрона, которое находится всегда вне головного и спинного мозга, располагаясь в узлах (ганглиях) периферической нервной системы. Затем по аксону нервный импульс направляется в центральную нервную систему, в спинной или в головной мозг. Поэтому чувствительные нейроны называют также приносящими (аффе-рентными) нервными клетками. Нервные окончания (рецепторы) различаются по своему строению, расположению и функциям. Выделяют экстеро-, интеро- и проприо-рецепторы. Экстерорецепторы воспринимают раздражение из внешней среды. Эти рецепторы находятся в наружных покровах тела (коже, слизистых оболочках), в органах чувств. Интерорецепторы получают раздражение в основном при изменении химического состава внутренней среды организма (хеморецепторы), давления в тканях и органах (барорецепторы). Проприо-рецепторы воспринимают раздражение (натяжение, напряжение) в мышцах, сухожилиях, связках, фасциях и суставных капсулах. В соответствии с функцией выделяют терморецеп-торы, которые воспринимают изменения температуры, и механорецепторы, улавливающие различные виды механических воздействий (прикосновение к коже, ее сдавление). Ноци-рецепторы воспринимают болевые раздражения.
Вставочные (ассоциативные, кондукторные) нейроны составляют до 97% нервных клеток нервной системы. Эти нейроны находятся, как правило, в пределах центральной нервной системы (головного и спинного мозга). Они передают полученный от чувствительного нейрона импульс эффекторному нейрону.
Эффекторные (выносящие или эфферентные) нейроны проводят нервные импульсы от мозга к рабочему органу — мышцам, железам и другим органам. Тела этих нейронов располагаются в головном и спинном мозге, в симпатических или парасимпатических узлах на периферии.
Нервные волокна представляют собой отростки нервных клеток (дендриты, аксоны), покрытые оболочками (рис. 295). При этом отросток в каждом нервном волокне является осевым цилиндром, а окружающие его нейролеммоциты (шванновские клетки), относящиеся к нейроглии, образуют оболочку волокна — нейролемму. С учетом строения оболочек нервные волокна подразделяют на безмякотные (безмиелиновые) и мякотные (миелиновые).
Безмиелиновые нервные волокна имеются, главным образом, у вегетативных нейронов. Осевой цилиндр как бы прогибает плазматическую мембрану (оболочку) нейролеммоцита, которая смыкается над ним. Сдвоенная над осевым цилиндром мембрана нейролеммоцита получила название мезаксон. Под шванновской клеткой остается узкое пространство (10-15 нм), содержащее тканевую жидкость, участвующую в проведении нервных импульсов. Один нейролеммоцит окутывает несколько (до 5-20) аксонов нервных клеток. Оболочку отростка нервной клетки образуют многие шванновские клетки, располагающиеся последовательно одна за другой.
Миелиновые нервные волокна толстые, они имеют толщину до 20 мкм. Эти волокна образованы довольно толстым аксоном клетки — осевым цилиндром. Вокруг аксона имеется оболочка, состоящая из двух слоев. Внутренний слой, миелиновый, образуется в результате спирального накручивания нейролеммоцита (шванновской клетки) на осевой цилиндр (аксон) нервной клетки. Цитоплазма нейролеммоцита выдавливается из него подобно тому, как это происходит при закручивании периферического конца тюбика с зубной пастой. Таким образом, миелин представляет собой многократно закрученный двойной слой плазматической мембраны (оболочки) нейролеммоцита. Толстая и плотная миелиновая оболочка, богатая жирами, изолирует нервное волокно и предотвращает утечку нервного импульса из аксолеммы (оболочки аксона).
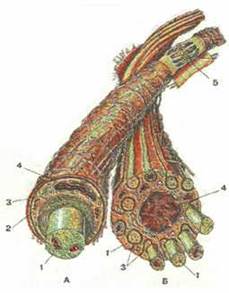
Рисунок 295. Нервные волокна. А — миелиновое волокно. Б — безмиелиновое волокно.
1 - осевой цилиндр; 2 - миелиновый слой; 3 - мезаксон; 4 - ядро нейролеммоцита (шванновской клетки); 5 - узловой перехват (перехват Ранвье).
Снаружи от миелинового находится тонкий слой, образованный самой цитоплазмой нейролеммоцитов. Дендриты миелиновой оболочки не имеют. Каждый нейролеммоцит (шванновская клетка) окутывает по длине только небольшой участок осевого цилиндра. Поэтому миелиновый слой не сплошной, прерывистый. Через каждые 0,3-1,5 мм имеются так называемые узловые перехваты нервного волокна (перехваты Ранвье), где миелиновый слой отсутствует. В этих местах соседние нейролеммоциты (шванновские клетки) своими концами подходят непосредственно к осевому цилиндру. Перехваты Ранвье способствует быстрому прохождению нервных импульсов по миелиновым нервным волокнам. Нервные импульсы по миелиновым волокнам проводятся как бы прыжками — от перехвата Ранвье к следующему перехвату.
Скорость проведения нервных импульсов по безмиелиновым волокнам составляет 1-2 м/с, а по мякотным (миелиновым) — 5-120 м/с. По мере удаления от тела нейрона скорость проведения импульса уменьшается.
Синапсы. Нейроны нервной системы вступают в контакт друг с другом и образуют цепочки (рис. 296), по которым передается нервный импульс.
Рисунок 296. Схема передачи нервного импульса.
Периферическая нервная система представлена аксонами нервных клеток (осевыми цилиндрами), которые либо покрыты неврилеммой шванновских клеток (безмякотные волокна), либо между неврилеммой и осевым цилиндром имеется многослойная миэлиновая оболочка (мякотные волокна). Мякотные и безмякотные волокна объединяются в пучки, ограниченные трубчатой соединительно-тканной оболочкой — периневрием. Внутри периневральной трубки каждое нервное волокно окружает рыхлая соединительная ткань (эндоневрий). Пучки волокон, покрытые оболочкой, называют нервами. Часто пучки переходят в более толстые образования — нервные стволы, в которых несколько пучков окружены рыхлой соединительной тканью — эпиневрием.
Нейроны соединяются между собой несколькими способами. Наиболее примитивным и древним является протоплазматический способ, когда отросток одной нервной клетки переходит в отросток другой клетки. Если нервные клетки контактируют между собой немиэлинизированными участками сомы или отростков и появляется возможность электро-тонического взаимодействия, соединение называют эфаптическим. Третий способ соединения между нейронами, а также нейрона с клетками, не принадлежащими к нервной системе (мышечными, желудочными), — синаптический — наиболее сложный. Он предполагает наличие специального структурного образования — синапса.
Синапсами называют специализированные контакты между клетками, используемые для передачи сигналов. Синапс состоит из окончания пресинаптического нейрона, постсинап-тической структуры и синаптической щели между ними. Пресинаптические терминали аксона расширяются, образуя концевую «пуговку» («бляшку»), окруженную аксолеммой. Ее участок, почти вплотную прилегающий к постсинаптической мембране другой клетки, называется пресинаптической мембраной. В цитоплазме синаптической бляшки много митохондрий и синаптических пузырьков (везикул) диаметром 40-50 нм.
Ширина синаптической щели в химических синапсах — 20-30 нм, а в электрических — 2-4 нм. Синапсы классифицируют по их расположению на поверхности постсинаптического нейрона. Если аксон оканчивается на дендрите другого нейрона, это аксо-дендритный синапс (рис. 297.1.) (часто синаптическая бляшка как бы «надевается» на специальные выступы — дендритные шипики), если же на соме другого нейрона, это аксо-соматический синапс (около половины поверхности сомы и почти вся поверхность дендритов может быть усеяна контактами от других нейронов). Аксон образует иногда синапсы в своей проксимальной части, лишенной миэлина, либо на синаптической бляшке другого нейрона. Такие синапсы являются аксо-аксонными (В, Д). Реже встречаются синапсы между дендритами (дендро-дендритные) (Б) и между дендритными шипиками и телом другого нейрона (дендро-сома-тические). В окончаниях периферических нервов на мышцах имеются нервно-мышечные (мионевральные) синапсы, на железах — нейросекреторные, а на внутренних органах — органные синапсы. Аксо-васкулярные синапсы наблюдают между нейросекреторными клетками гипоталамуса и стенками капилляров.
Синапсы можно классифицировать, во-первых, по их местоположению и принадлежности соответствующим клеткам (нервно-мышечные, нейро-нейрональные, аксо-соматические, аксо-дендритические и т.д.). Во-вторых, синапсы можно разделить по знаку их действия на возбуждающие и тормозящие. И, наконец, по способу передачи сигналов они разделяются на электрические, в которых сигналы передаются электрическим током, и химические, в которых передатчиком сигнала (трансмиттер) или иначе посредником (медиатор) является то или иное физиологически активное вещество. Существуют и смешанные — электрохимические синапсы. Заметим, что и в том, и в другом синапсе имеются такие компоненты, как пресинаптическая мембрана, постсинаптическая мембрана и разделяющая их синаптическая щель.
В мозге редко встречаются изолированные одиночные синапсы. Обычно несколько синапсов вместе складываются в тот или иной тип групповой синаптической связи. Простейший из таких типов — когда два или несколько синапсов расположены рядом друг с другом и ориенти-рованы в одном направлении; все они бывают аксо-дендритными. Более сложен тип, в котором отросток а образует синапс на отростке б, а отросток б на отростке в. Такая ситуация схематически показана на рис. 297.1.Д. Такие синапсы называют последовательными; их примерами могут служить аксо-аксодендритные и аксо-дендродендритные последовательности.
Еще в одном типе отросток а соединяется с отростком б, а последний — снова с отростком а. Эта ситуация схематически показана на рис. 297.1.Г. Такой синапс принято называть реципрокным. Если два таких синапса расположены рядом, то их называют реципрокной парой. Если же два синапса удалены один от другого, то возникает реципрокное устройство. Наконец, есть такие типы синаптических соединений, когда тесно сближена целая группа терминалей. Этот тип называют синаптической гломерулой (рис. 297.1.Е).
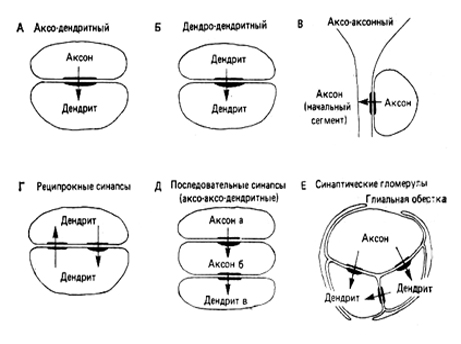
Рисунок 297. Виды синапсов.
Передача информации в синапсе осуществляется специальными химическими веществами-посредниками (медиаторами), выделяемыми из нервных окончаний в синаптическую щель. В нервной системе эти вещества называют нейромедиаторами. Основными нейромедиаторами в вегетативной нервной системе являются ацетилхолин и норадреналин. В состоянии покоя эти медиаторы, вырабатываемые в нервных окончаниях, находятся в особых пузырьках.
Работа этих медиаторов показана на рисунке 298. Условно (так как он занимает считанные доли секунды) весь процесс передачи информации можно разбить на четыре этапа. Как только по пресинаптическому окончанию поступает импульс, на внутренней стороне клеточной мембраны за счет входа ионов натрия происходит образование положительного заряда, и пузырьки с медиатором начинают приближаться к пресинаптической мембране (этап I). На втором этапе осуществляется выход медиатора в синаптическую щель из пузырьков в месте их контакта с пресинаптической мембраной. После выделения из нервных окончаний нейромедиатор проходит синаптическую щель путем диффузии и связывается со своими рецепторами постсинаптической мембраны клетки исполнительного органа или другой нервной клетки (этап III). Активация рецепторов запускает в клетке биохимические процессы, приводящие к изменению ее функционального состояния в соответствии с тем, какой сигнал был получен от афферентных звеньев. На уровне органов это проявляется сокращением или расслаблением гладких мышц (сужением или расширением сосудов, учащением или замедлением и усилением или ослаблением сокращений сердца), выделением секрета и так далее. И, наконец, на четвертом этапе происходит возвращение синапса в состояние покоя либо за счет разрушения медиатора ферментами в синаптической щели, либо благодаря транспорту его обратно в пресинаптическое окончание. Сигналом к прекращению выделения медиатора служит возбуждение им рецепторов пресинаптической мембраны.
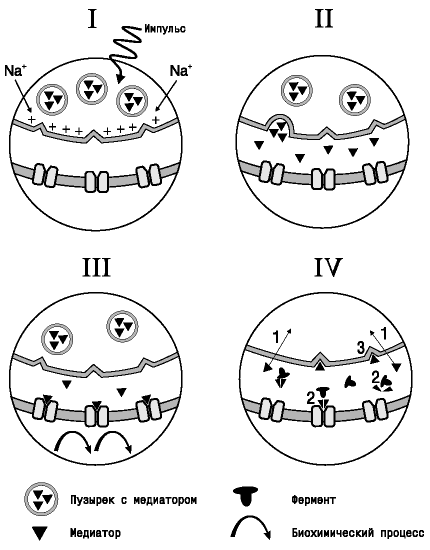
Рисунок 298. Функционирование синапса: I – поступление нервного импульса; II – выделение медиатора в синаптическую щель; III – взаимодействие с рецептором постсинаптической мембраны; IV – «судьба» медиатора в синаптической щели – возвращение синапса в состояние покоя. 1 – обратный захват медиатора; 2 – разрушение медиатора ферментом; 3 – возбуждение пресинаптических рецепторов.
Выше уже сказано, что в вегетативной нервной системе передача информации осущест-вляется, главным образом, с помощью медиаторов — ацетилхолина и норадреналина. Поэтому пути передачи и синапсы называют холинергическими (медиатор — ацетилхолин) или адренергическими (медиатор — норадреналин). Аналогично этому рецепторы, с кото-рыми связывается ацетилхолин, называют холинорецепторами, а рецепторы норадреналина — адренорецепторами. На адренорецепторы влияет также гормон, выделяемый надпочеч-никами, — адреналин.
Холино- и адренорецепторы неоднородны и различаются чувствительностью к некоторым химическим веществам. Так, среди холинорецепторов выделяют мускаринчувствительные (м-холинорецепторы) и никотинчувствительные (н-холинорецепторы) — по названиям естественных алкалоидов, которые оказывают избирательное действие на соответствующие холинорецепторы. Мускариновые холинорецепторы, в свою очередь, могут быть м1-, м2- и м3-типа в зависимости от того, в каких органах или тканях они преобладают. Адренорецепторы, исходя из различной чувствительности их к химическим соединениям, подразделяют на альфа- и бета-адренорецепторы, которые тоже в зависимости от локализации имеют несколько разновидностей.
Сеть нервных волокон пронизывает все человеческое тело, таким образом, холино- и адренорецепторы расположены по всему телу. Нервный импульс, распространяющийся по всей нервной сети или ее пучку, воспринимается как сигнал к действию теми клетками, которые имеют соответствующие рецепторы. И, хотя холинорецепторы локализуются в большей степени в мышцах внутренних органов (желудочно-кишечного тракта, мочеполовой системы, глаз, сердца, бронхиол и других органов), а адренорецепторы — в сердце, сосудах, бронхах, печени, почках и в жировых клетках, обнаружить их можно практически в каждом органе. Воздействия, при реализации которых они служат посредниками, очень разно-образны.
Зная механизм передачи информации в вегетативной нервной системе, можно предположить, как и в каких местах этой передачи нам необходимо действовать, чтобы выз-вать определенные эффекты. Для этого мы можем использовать вещества, которые имити-руют (миметики) или блокируют (литики) работу нейромедиаторов, угнетают действие ферментов, разрушающих эти медиаторы, или препятствуют высвобождению посредников из пресинаптических пузырьков. Используя такие лекарства, можно оказывать влияние на многие органы: регулировать деятельность сердечной мышцы, желудка, бронхов, стенок сосудов и так далее.
В ответ на раздражение нервная ткань приходит в состояние возбуждения, которое представляет собой нервный процесс, вызывающий или усиливающий деятельность органа. Свойство нервной ткани передавать возбуждение называется проводимостью. Скорость проведения возбуждения значительна: от 0,5 до 100 м/с, поэтому между органами и системами быстро устанавливается взаимодействие, отвечающее потребностям организма. Возбуждение проводится по нервным волокнам изолированно и не переходит с одного волокна на другое, чему препятствуют оболочки, покрывающие нервные волокна.
Клетки нейроглии в нервной системе подразделяются на два вида. Это глиоциты (или макроглия) и микроглия.
Среди глиоцитов различают эпендимоциты, астроциты и олигодендроциты.
Эпендимоциты образуют плотный слой, выстилающий центральный канал спинного мозга и все желудочки головного мозга. Они участвуют в образовании спинномозговой жидкости, транспортных процессах, в метаболизме мозга, выполняют опорную и разграничительную функции. Эти клетки имеют кубическую или призматическую форму, располагаются они в один слой. Их поверхность покрыта микроворсинками.
Астроциты образуют опорный аппарат центральной нервной системы. Они представляют собой мелкие клетки с многочисленными, расходящимися во все стороны отростками. Различают волокнистые и протоплазматические астроциты. Волокнистые астроциты имеют 20-40 длинных, слабо ветвящихся отростков, преобладают в белом веществе центральной нервной системы. Отростки располагаются между нервными волокнами. Некоторые отростки достигают кровеносных капилляров. Протоплазматические астроциты располагаются преимущественно в сером веществе центральной нервной системы, имеют звездчатую форму, от их тел во все стороны отходят короткие сильно разветвленные, многочисленные отростки. Отростки астроцитов служат опорой для отростков нейронов, образуют сеть, в ячейках которой залегают нейроны. Отростки астроцитов, достигающие поверхности мозга, соединяются между собой и образуют на ней сплошную поверхностную пограничную мембрану.
Олигодендриты — наиболее многочисленная группа клеток нейроглии. Они окружают тела нейронов в центральной и периферической нервной системе, находятся в составе оболочек нервных волокон и нервных окончаний. Олигрдендроциты представляют собой мелкие овоидные клетки диаметром 6-8 мкм с крупным ядром. Клетки имеют небольшое количество отростков конусовидной и трапециевидной формы. Отростки образуют миелиновый слой нервных волокон. Миелинообразующие отростки спирально накручиваются на аксоны. По ходу аксона миелиновая оболочка сформирована отростками многих олигодендроцитов, каждый из которых образует один сегмент. Между сегментами находится лишенный миелина узловой перехват нервного волокна (перехват Ранвье). Олигодендроциты, образующие оболочки нервных волокон периферической нервной системы, называются нейролеммоцитами (шванновскими клетками).
Микроглия составляет около 5% клеток нейроглии в белом веществе мозга и 18% в сером веществе. Микроглия представлена мелкими удлиненными клетками угловатой или неправильной формы, рассеянными в белом и сером веществе (клетки Ортега). От тела каждой клетки отходят многочисленные отростки разной формы, напоминающие кустики, которые заканчиваются на кровеносных капиллярах. Ядра клеток имеют вытянутую или треугольную форму. Микроглиоциты обладают подвижностью и фагоцитарной способностью. Они выполняют функцию своеобразных «чистильщиков», поглощая частицы погибших клеток.
На срезах ЦНС видны участки серого и белого цветов. Это серое и белое веще
| <== предыдущая лекция | | | следующая лекция ==> |
| Возникновение, сущность и функции денег. Современные виды денег | | | Превращение денег в капитал. Рабочая сила как товар |
Дата добавления: 2016-07-27; просмотров: 2082;
Поиск по сайту
Узнать еще
- II пара – зрительный нерв, nervus opticus.
- II. Абсолютное пространство по самой своей сущности, безотносительно к чему бы то ни было внешнему, остается всегда одинаковым и неподвижным».
- IX пара – языкоглоточный нерв, nervus glossopharyngeus
- благодаря чему переадресация может быть выполнена в любой момент: до начала монтажных работ, в процессе их осущствления и после ввода системы в эксплуатацию.
- В связи с большим количеством подземных коммуникаций и инженерных сооружений геодезические работы выполняют в городских условиях с большей точностью, чем на загородных дорогах.
- В чем же заключается проблема формы в учебном рисунке?
- В чем же заключается экстренная помощь при ожоговых травмах?
- В чем же, однако, сама проблема?

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
