ГЕНЕТИКА И ГЕНЕТИЧЕСКИЕ МЕТОДЫ ИДЕНТИФИКАЦИИ МИКРООРГАНИЗМОВ
Цель занятия. Ознакомить студентов с фенотипической и генотипической изменчивостью и генетическими методами идентификации микроорганизмов.
Оборудование и материалы. Культура Proteus vulgarus, пробирки с 2%-м МПА, 1%-й раствор фенола, пробирки с 10 мл и с 1 мл МПБ, чашки Петри с 3%-м МПА, содержащим 100 ЕД стрептомицина в 2 мл среды, чашки Петри с обычным МПА, стерильные мерные пипетки на 1 мл, диссоциирующая культура Е. coli или P. multocida на МПА в чашках Петри в виде изолированных колоний, бинокулярная лупа МБС-1, культура Е. coli, продуцирующая колицин, и колицинчувствительная культура Е. coli, чашки Петри с 1,5%-м МПА, хлороформ, пробирки с 0,7%-м МПА, штаммы Е. coli K12F-, lew, strr, Е. coli К12 Hfr, leu+, strs, E. coli lac-, трансдуцирующий фаг λ dgal, содержащий β-галактозидазный оперон Е. coli; набор компонентов для ПЦР-диагностики какой-либо инфекционной болезни, амплификатор.
МЕТОДИЧЕСКИЕ УКАЗАНИЯ
Генетика микроорганизмов. Природа изменений свойств микроорганизмов может быть наследственной (генотипической) и ненаследственной (модификационной, фенотипической).
Модификационная изменчивость. В отличие от мутаций затрагивает сразу большое количество клеток в популяции.
Модификации — это ненаследуемые фенотипические изменения, которые возникают под влиянием различных факторов, но после прекращения их действия исчезают. При этом клетки с одинаковым генотипом могут различаться по фенотипу.
Генотипическая изменчивость. Может быть мутационной, обусловленной изменениями в ДНК под влиянием мутагенов, и комбинативной, связанной с процессом образования новых комбинаций генов в генотипе.
Мутации проявляются фенотипически в виде изменения какого-либо признака, присущего микроорганизму, — физиологического, морфологического. Различают индуцированные и спонтанные мутации. Последние возникают под влиянием неустановленных факторов и достаточно редки в популяции (1 • 10-5...1*10-7).
Рекомбинации — обмен генетическим материалом между двумя бактериальными клетками. Клетку, воспринимающую генетический материал, называют реципиентом, отдающую — донором. Рекомбинантные клетки содержат в составе своего генома фрагменты хромосомы донора. Механизм проникновения донорской ДНК в клетку реципиента может быть различен: 1) трансформация — «голая» ДНК проникает в клетку реципиента без взаимного контакта клеток; 2) трансдукция — перенос фрагментов ДНК из клетки донора в реципиентную клетку умеренным бактериофагом; 3) конъюгация — перенос фрагмента ДНК при непосредственном контакте клеток донора и реципиента. В процессе конъюгации из одной клетки в другую может переходить плазмидная ДНК. Плазмида (F-фактор) представляет собой внехромосомные небольшие кольцевые молекулы ДНК разной длины. Плазмиды обычно обозначают по характерному свойству клетки-хозяина: R-фактор — плазмида, обеспечивающая устойчивость к лекарственным препаратам, Соl-фактор — плазмида, детерминирующая синтез клеткой антибиотикоподоб-ных веществ (колицинов, бактериоцинов) и т. д.
Выявление модификационной изменчивости. В пробирку с 6 мл расплавленного МПА добавляют 0,6 мл 1%-го раствора фенола, агар скашивают. В пробирки с фенолизированным и обычным МПА засевают культуру Proteus vulgaris по методу Шукевича в конденсационную воду. Посевы инкубируют при 37...38 °С 24 ч и учитывают характер роста протея на фенолизированном и обычном МПА. На фенолизированном агаре протей в отличие от обычного МПА не проявляет феномена «роения».
Выявление мутационной изменчивости во флуктуационном тесте Лурия—Дельбрюка. Данный тест используют для изучения природы возникновения различных свойств у бактерий: устойчивости к бактериофагу, лекарственным препаратам, способности расщеплять углеводы и т. д. В соответствии с гипотезой адаптации каждая клетка в популяции в присутствии того или иного фактора — фага, антибиотика и т. д. — может с некоторой долей вероятности приобрести устойчивость к нему (адаптироваться). Согласно гипотезе спонтанных мутаций каждая клетка бактерий также с небольшой долей вероятности может приобрести устойчивость к тому или иному фактору в результате мутаций независимо от присутствия этого фактора в среде обитания.
С помощью флуктуационного теста Лурия—Дельбрюка можно подтвердить мутационную природу возникшего свойства.
Готовят два ряда культур.
1. «Параллельные» культуры: в пробирку, содержащую 10 мл МПБ (обозначим ее № 1), и в 10 пробирок, содержащих по 1 мл МПБ (№2...11), одновременно высевают одинаковое количество клеток Е. coli (300...500 бактериальных клеток на пробирку). Посевы инкубируют при 37 °С 24 ч.
2. «Независимые» культуры: в 20 чашек Петри разливают МПА с добавлением стрептомицина (конечная концентрация антибиотика 100 ЕД на чашку). Первые 10 чашек засевают по 0,1 мл культурой, взятой из пробирки № 1; следующие 10 чашек засевают по 0,1мл культурами, взятыми из пробирок №2...11. Посевы инкубируют при 37 ºС 24 ч. После инкубирования подсчитывают число выросших колоний на всех чашках Петри с «параллельными» и «независимыми» культурами.
На стрептомициновом агаре давать колонии могут только устойчивые к стрептомицину клетки Е. coli.
Примерно одинаковое число колоний на всех чашках (нет статистически достоверных отличий) подтверждает теорию адаптации. Если же на чашках, засеянных культурами из пробирок №2...11, наблюдают существенную разницу в числе колоний, а на чашках, засеянных культурой из пробирки № 1, нет статистически достоверных отличий, то можно говорить о мутационной природе наблюдаемой стрептомицинустойчивости. Мутации — редкое событие, и при посеве из отдельных пробирок мутанты вырастают только в некоторых чашках. При посеве же из общей пробирки предсуществующие мутанты будут равномерно распределены в жидкой среде и соответственно в чашках с агаром.
Диссоциация бактерий.Диссоциация — это появление в бактериальной популяции колоний различного типа: S-формы (smooth — гладкий), R-формы (rough — шероховатый), что может быть следствием мутаций или рекомбинаций.
Для бактерий в S-форме характерны типичные для вида морфологические, ферментативные и вирулентные свойства. У R-формы обычно отмечают меньшую вирулентность, потерю способности к капсулообразованию, нередко изменение ферментативных свойств. Анализ популяции бактериальных штаммов по признаку диссоциации с целью отвивки (селекции) S-формы — обязательный этап при работе с производственными штаммами. В ходе такой селекции ориентируются на морфологические и оптические характеристики S- и R-колоний.
Для анализа популяции бактерий с признаками диссоциации (Е. coli, P. multocida) производят дробный рассев исследуемой культуры на МПА в чашки Петри с целью получения изолированных колоний. Посевы культивируют при 37...38 °С 18...24 ч. Чашку Петри с колониями помещают на предметный столик бинокулярной лупы и изучают в косопроходящем пучке света (см. тему 8).
Дополнительно из колоний S- и R-формы делают окрашенные мазки и изучают особенности морфологии клеток.
Выявление бактериальной плазмиды Со1+ (по методу отсроченного антагонизма Фредерик). Исследуемую культуру Е. coli засевают уколом в 1,5%-й МПА (обычно 6...8 штаммов на одну чашку). Посевы инкубируют при 37 °С 48 ч. Выросшие колонии убивают парами хлороформа, затем поверхность агара заливают слоем расплавленного и охлажденного до 46...48 ºС 0,7%-го агара, смешанного с 0,1 мл 6-часовой бульонной культуры Е. coli, чувствительной к колицинам. Через 24 ч инкубирования при 37 ºС вокруг колоний, продуцирующих колицин, можно видеть зоны подавления роста колицинчувствительной (индикаторной) культуры.
Демонстрация специфической трансдукции. Для проведения опыта необходимы: реципиентный штамм Е. coli lac-, не способный ферментировать лактозу, трансдуцирующий фаг λ dgal, геном которого содержит β-галактозидазный оперон Е. coli.
Смешивают 1 мл 3-часовой бульонной культуры Е. coli и 1 мл фага с титром 10-6...10-7, смесь инкубируют при 37 °С 1 ч. Затем из смеси готовят десятикратные разведения в пробирках. Из пробирки с разведением 10-6 высевают в три чашки Петри с агаром Эндо по 0,1мл культуры. Посевы инкубируют при 37 ºС 24 ч. Подсчитывают лактозонегативные и лактозопозитивные (транс -дуктанты) колонии. Соотносят число трансдуктантов к числу колоний реципиентного штамма (светлые, лактозонегативные колонии) и определяют таким образом частоту трансдукции. Например, выросло 6 колоний красного цвета (трансдуктанты) и 468 светлых колоний (реципиентные), следовательно, частота трансдукции составляет 6/468 = 1,3 * 10-2.
Демонстрация конъюгации. Для проведения опыта необходимы: реципиентный штамм Е. coli K12F-, leu-, Strr; донорский штамм Е. coli К12, Hfr, leu+, Strs; минимальная синтетическая среда для обнаружения рекомбинантов (сульфат железа — 0,0005 г, глюкоза — 2 г, сульфат аммония — 1 г, сульфат магния — 1 г, ди-гидрофосфат калия —6,5 г, нитрат кальция — 0,001 г, стрептомицин — 200 ЕД/мл, вода дистиллированная — 1000 мл), аналогичная питательная среда без стрептомицина; полноценный питательный агар со стрептомицином.
В пробирке объединяют 3-часовые бульонные культуры реципиентного и донорского штаммов в соотношении 2:1, смесь выдерживают при 37 °С 30 мин, разводят стерильным физиологическим раствором в соотношении 1 : 100 и по 0,1 мл высевают на минимальную селективную среду (агаровую). Посевы инкубируют при 37 °С 18...20 ч, затем учитывают рост колоний эшерихий. На этой среде могут расти только клетки рекомбинантов, соединившие в себе устойчивость к стрептомицину (свойство реципиента) и отсутствие зависимости от лейцина (свойство донора). Для отрицательного контроля на минимальную селективную среду высевают культуры донорского и реципиентного штаммов (не должны расти). Для положительного контроля донорский штамм высевают на селективную среду без стрептомицина, а реципиентный штамм — на полноценный питательный агар со стрептомицином (эшерихий должны расти на обеих средах).
Если определить количество жизнеспособных клеток реципиентов в посевном материале и соотнести их с количеством рекомбинантных клеток, то можно рассчитать частоту рекомбинации (см. «Демонстрацию специфической трансдукции»).
Генетические методы идентификации микроорганизмов. Фенотипические признаки бактерий отражают максимум 20 % генотипа, поэтому ученые интенсивно разрабатывают методы и критерии идентификации микроорганизмов, основанные на изучении не фенотипического проявления генома, а особенностей строения ДНК.
Сравнение геномов по составу основания ДНК.ДНК содержит четыре азотистых основания: аденин (А), тимин (Т), гуанин (Г), цитозин (Ц). По правилам спаривания оснований А = Т и Г = Ц, но молярное отношение (Г + Ц): (А + Т) варьирует у разных видов и может служить таксономической характеристикой.
Поскольку число водородных связей между основаниями в парах ГЦ больше, чем между основаниями в парах AT, то о ГЦ-содержании судят по «температуре плавления» ДНК, т. е. по температуре, при которой происходит разрыв водородных связей, соединяющих цепи ДНК. Разделению цепей сопутствует увеличение оптической плотности, которую измеряют спектрофотометром при А = 260 нм (максимум поглощения ДНК). По мере нагревания образца ДНК поглощение возрастает и, когда ДНК становится одноцепочечной, кривая поглощения достигает плато.
У каждого бактериального вида ДНК содержит характерное среднее количество ГЦ, и если нуклеотидный состав ДНК у двух сравниваемых микроорганизмов существенно отличается(10...15 %), то говорят об их принадлежности к различным таксономическим группам. Однако отсутствия существенных отличий в количестве ГЦ у двух видов бактерий недостаточно для заключения о тождестве этих бактерий: необходимо доказать высокое фенотипическое сходство обоих видов или применить другие генетические методы.
Метод гибридизации нуклеиновых кислот. Основан на том, что одноцепочечная ДНК после тепловой денатурации при понижении температуры на 20...30 °С ниже температуры плавления способна реассоциироваться (отжиг) в изначальную двухцепочечную ДНК. Также был установлен факт реассоциации одноцепочечных ДНК от различных видов бактерий с образованием гибридной ДНК, причем чем больше гомология сравниваемых одноцепочечных ДНК, тем успешнее реассоциация. Аналогичным образом можно проводить гибридизацию между РНК и ДНК, тРНК и рРНК.
При проведении гибридизации ДНК один из сравниваемых образцов денатурированной ДНК метят радиоактивным изотопом, образовавшиеся двухцепочечные ДНК отделяют от одноцепочечных и измеряют их радиоактивность. В качестве контроля используют меченую и немеченую ДНК эталонного (известного) штамма микроорганизма. Степень реассоциации таких гомологичных одноцепочечных ДНК принимают условно за 100%. Существует точка зрения, что при 60... 100 % гомологии ДНК можно говорить о принадлежности сравниваемых бактерий к одному виду, при 40...60 % — к одному роду, при 0...25 % гомологии — к родам одного семейства и разным семействам.
Метод генных зондов. Используют достаточно часто для идентификации бактерий. От обычной ДНК—ДНК гибридизации этот способ отличается использованием не тотальной ДНК, а определенного ее фрагмента (зонда), содержащего известный ген (генетический маркер). Предварительно создают «банк генов» изучаемой бактерии. С этой целью бактериальную ДНК расщепляют эндонуклеазами, фрагменты ДНК разделяют электрофоретически, методом трансформации определяют их генетические характеристики, отбирают нужный фрагмент ДНК и при помощи лигазы включают в плазмиду, которая служит вектором. Плазмиду с интегрированным известным геном вводят в какой-либо бактериальный штамм, обычно достаточно легко культивируемый. Получают большое количество биомассы, содержащей ДНК-зонд. Выделяют плазмидную ДНК, метят ее радиоактивным изотопом, затем эту меченую ДНК гибридизируют с ДНК изучаемой бактерии. Методом ауторадиографии выявляют относительную частоту гибридизации маркера с изучаемой ДНК и по этому показателю судят о генетическом родстве известной (бактерии — донора ДНК) и исследуемой бактерий.
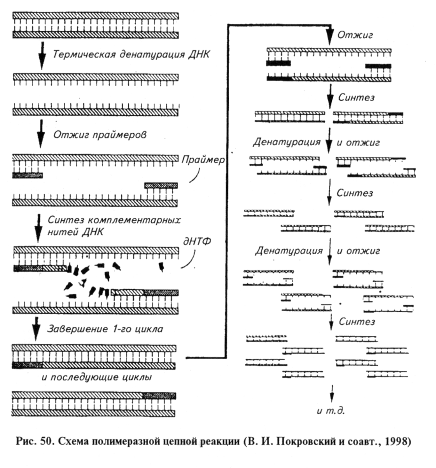
Полимеразная цепная реакция — ПЦР (polymerase chain reaction — PCR). Принцип реакции состоит в том, что при помощи ДНК-полимеразы in vitro многократно синтезируют копии определенного участка ДНК (амплификация — накопление).
ПЦР — циклический процесс, каждый цикл которого включает в себя три стадии.
1. Проводят тепловую денатурацию (95 °С) исследуемой ДНК. При этом разрушаются водородные связи, соединяющие парные основания, и цепи ДНК расходятся, т. е. образуются одноцепо-чечные ДНК, которые доступны для праймеров (затравок) и ДНК-полимеразы. Длительность процесса 1 мин.
2. Осуществляют посадку (отжиг) праймеров на комплементарные участки двух антипараллельных цепей ДНК. Праймеры представляют собой два синтетических олигонуклеотида длиной в 20...30 нуклеотидов, каждый из которых комплементарен противоположной цепи ДНК в участках, ограничивающих выбранный сегмент ДНК возбудителя. Таким образом, праймеры ограничивают специфический для возбудителя участок ДНК. Праймеры добавляют в реакционную смесь в избытке, что позволяет им занять свои комплементарные участки до того, как произойдет соединение (ренатурация) одноцепочечных ДНК в двухцепо-чечную. Длительность стадии 1...2мин.
3. Идет процесс достройки (элонгации) праймера из внесенных в реакционную смесь дезоксинуклеозидтрифосфатов при участии ДНК-полимеразы. Используют обычно термостабильную ДНК-полимеразу термофильной бактерии Thermus aquaticus (Taq-полимераза), что позволяет вести полимеризацию при оптимальной температуре 70...75 °С. При синтезе ДНК праймеры включаются в ее молекулу. Синтез ДНК с помощью полимеразы протекает только между праймерами, и при этом удваивается число копий именно этого участка ДНК. Молекула ДНК, синтезированная при помощи одного праймера, может служить матрицей для синтеза комплементарной ДНК при помощи другого праймера. Длительность этой стадии 1...2 мин (рис. 50).
По завершении 1-го цикла реакцию тормозят и ДНК вновь подвергают температурной денатурации. При охлаждении избыточные праймеры опять гибридизируются с цепями исходной и вновь синтезированной ДНК. Добавление ДНК-полимеразы обеспечивает второй цикл полимеризации. Таким образом можно провести несколько десятков циклов ферментативного удлинения праймеров, в результате количество сегментов ДНК, ограниченных с обеих сторон праймерами, с каждым циклом увеличивается экспоненциально. Следовательно, используя метод ПЦР, можно in vitro избирательно обогащать препарат ДНК фрагментом с определенной последовательностью в миллион раз и более. Факт увеличения числа фрагментов ДНК служит доказательством присутствия в исследуемом образце гомологичной ДНК, т. е. возбудителя инфекционной болезни.
Для практического использования ПЦР необходимо синтезировать праймеры, комплементарные З'-концам цепей копируемой ДНК-матрицы и ограничивающие копируемый фрагмент ДНК. Их выбирают на основе участков генома возбудителя с расшифрованной нуклеотидной последовательностью и устойчивых к генетическим перестройкам. Например, для идентификации C.psittaci были предложены праймеры, выбранные на основе гена, кодирующего белок наружной мембраны, или на основе гена, кодирующего 16s рРНК; для идентификации бруцелл был выбран праймер на основе гена, кодирующего белок внешней мембраны 31 КДа, и т. д.
Для проведения ПЦР-диагностики необходимы следующие компоненты: водные растворы дезоксинуклеозидтрифосфатов четырех типов (gATO, gTTO, glTO, gHTO, 10 мМ, рН 7,0); первый праймер (5 мМ); второй праймер (5 мМ); фермент Taq-поли-мераза (5 ЕД/мкл); амплифицируемая ДНК (~ 1 мкг); ионы Mg2+ для обеспечения работы полимеразы (25 мМ); буферный раствор (десятикратный концентрат), например, трис-соляная кислота (рН 6,8...7,8) с добавлением бычьего альбумина и неионных детергентов.
Указанные компоненты объединяют в пробирке, например, в следующих количественных соотношениях: раствор дезоксинуклеозидтрифосфатов — 8 мкл, буфер—10мкл, амплифицируемая ДНК — ~ 1 мкг, праймеры — по 1...5 мкл, полимераза — 0,5 мкл, дистиллированная вода — до 100 мкл. Для предотвращения испарения на жидкость в пробирке наслаивают минеральное масло. На первом этапе проведения ПЦР денатурируют ДНК, затем в реакционную смесь, содержащую дезоксинуклеозидтрифосфаты, вносят полимеразу и помещают в амплификатор или термоцик-лер (программируемый термостат). Амплификацию проводят в термоциклере по заданной программе, например: 90 "С — 1 мин; 60 °С - 1 мин; 72ºС - 1 мин (5 циклов); 93 °С - 1 мин; 57 °С -1 мин; 92 °С — 1 мин (5 циклов); 93 °С — 1 мин; 55 °С — 1 мин; ,72 °С — 3 мин (25 циклов).
После завершения амплификации следует этап обнаружения продуктов ПЦР — амплификонов. Молекулы ДНК и их фрагменты разделяют электрофорезом в агаровом геле. ДНК в геле окрашивают бромидом этидия с последующим анализом и фотографированием фореграмм при ультрафиолетовом облучении. Специфичность полосы амплифицированной ДНК подтверждают сравнением с маркерными фрагментами и ДНК-стандартом. Дополнительно специфичность амплификона можно подтвердить путем гибридизации со специфическим радиоактивным зондом.
Объектом исследования в ПЦР могут служить как ткани, содержащие возбудитель, так и культуры возбудителя. Из материала обычно тем или иным способом извлекают ДНК. В зависимости от характера материала методы его обработки варьируют.
Ценность ПЦР-диагностики возрастает, когда необходимо обнаружить возбудителей инфекционных болезней, или трудно культивируемых, или находящихся в нетипичной форме (L-формы бактерий), а также выявить гены, контролирующие тот или иной фактор патогенности микроорганизма.
ЗАДАНИЯ ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ
1. Изучить и охарактеризовать рост культуры протея на обычном и фенолизированном МПА.
2. Определить природу возникновения стрептомицинрезистентных форм Е. coli по результатам флуктуационного теста Лурия—Дельбрюка.
3. Ознакомиться с диссоциацией бактерий. Описать особенности морфологии S- и R-колоний и бактериальных клеток из них.
4. Ознакомиться с бактериоциногенией (способность выделять антибиотикоподобные вещества) на примере колицинов Е. coli.
5. Проанализировать результаты опытов по конъюгации,
трансдукции и трансформации бактерий.
6. Изучить схему проведения ПЦР, реагенты, аппаратуру.
Контрольные вопросы
1.Какие известны формы изменчивости бактерий?
2.В чем состоит тест Лурия—Дельбрюка?
3.Какова роль плазмид в формировании патогенных свойств бактерий?
4.Какие генотипические методы применяют для идентификации бактерий?
Раздел II
ИНФЕКЦИЯ И МЕТОДЫ ПРИКЛАДНОЙ ИММУНОЛОГИИ
•
Тема 13
Дата добавления: 2016-07-22; просмотров: 2834;
Поиск по сайту
Узнать еще
- I. История открытия и методы исследования вирусов
- I.2. Антигены системы АВ0. Генетика. Структура
- II. Категории и методы политологии.
- III. Методы искусственной физико-химической детоксикации.
- Імунологія і генетика туберкульозу
- Абсолютный возраст горных пород и методы его определения
- Автогенетические процессы и половой отбор
- Автоматические методы изготовления фотошаблонов.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
