Мегаспорогенез и мегагаметогенез
Мегаспоры образуются в процессе мегаспорогенеза в нуцеллусе (мегаспорангий) семязачатка (рис. 132). Обычно в субэпидермальном слое нуцеллуса вблизи верхушки семязачатка (в области микропиле) появляется одна или несколько клеток археспория. Она (если археспорий одноклеточный) и становится непосредственно материнской клеткой мегаспор. Как и все растение, материнская клетка мегаспор имеет диплоидный набор хромосом. Затем в результате мейоза из нее возникает тетрада гаплоидных мегаспор. Таким образом происходит мегаспорогенез. Чаще мегаспоры располагаются линейно от микропиле к халазе.
После этого начинается мегагаметогенез, т. е. формирование женского гаметофита — зародышевого мешка. Одна из мегаспор, обычно нижняя, находящаяся ближе к халазе, делится быстрее, в результате чего развитие остальных трех мегаспор подавляется: они сплющиваются, дегенерируют и рассасываются.
Прорастание мегаспоры и развитие женского гаметофита начинаются с разрастания ее клетки и трехкратного митотического деления ядра мегаспоры (первичного ядра зародышевого мешка). В конечном итоге в сильно растянутой клетке мегаспоры формируются восемь ядер, из которых четыре располагаются на ее микропилярном полюсе, четыре — на халазальном. Между ядрами происходит некоторая дифференцировка. С каждого полюса проросшей мегаспоры к ее центру отходит по одному ядру. Эти ядра называются полярными. Оставшиеся ядра обособляются в клетки. Одна из клеток на микропилярном полюсе отличается большим размером и преобразуется в яйцеклетку. Две рядом расположенные одинаковые клетки (синергиды) являются вспомогательными. Яйцеклетка вместе с синергидами образует яйцевой аппарат. Оставшиеся на хазальном (противоположном микропилярному) полюсе три клетки также обособляются. Эти клетки составляют группу антипод. В центре два полярных ядра сливаются, образуя вторичное (центральное) ядро зародышевого мешка.
В таком состоянии зародышевого мешка яйцеклетка и вторичное ядро готовы к оплодотворению.
Онтогенез цветка
8.10.1. Цветение. Сущность цветения состоит во вскрывании пыльников и функционировании рылец пестиков как органов, воспринимающих пыльцу. Цветение начинается с распускания цветков, т. е. с перехода от бутонизации к раскрытому околоцветнику. Распускание особенно заметно у цветков с ярким венчиком.
После опыления цветение завершается. Конец цветения может сопровождаться опадением частей цветков (чашелистиков, лепестков, тычинок), целых цветков или соцветий.
8.10.2. Монокарпия и поликарпия. Растения, которые цветут и плодоносят один раз в жизни, после чего погибают, называют монокарпическими или монокарпиками. К ним относятся все однолетние (лен, конопля, рожь, пшеница) и двулетние (редька, редис, капуста, свекла, морковь) растения.
У большинства многолетних растений цветение и плодоношение многократно повторяются в течение жизни. Их называют поликарпическими или поликарпиками. Лишь немногие многолетние растения являются монокарпиками, т. е. после первого цветения и плодоношения отмирают (некоторые агавы, бамбуки, пальмы, ферулы).
8.10.3. Формула и диаграмма цветка. Строение цветка можно выразить в виде формулы. При ее составлении пользуются следующими обозначениями: чашечка (Calyx) — Са, венчик (Corolla) — Со, андроцей (Androeceum) — А, гинецей (Gynoeceum) — G, простой околоцветник (Perigonium) — Р.
Типы цветков имеют также условные обозначения: обоеполый - 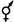 (этот значок в формуле обычно опускают), пестичный -
(этот значок в формуле обычно опускают), пестичный - 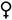 , тычиночный -
, тычиночный - 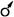 , актиноморфный - *, зигоморфный - , или │, асимметричный
, актиноморфный - *, зигоморфный - , или │, асимметричный 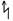 ,
, 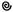 - цветок спиральный,
- цветок спиральный,  - цветок циклический.
- цветок циклический.
Число членов каждой части цветка обозначают цифрами (пятилепестный венчик — Со5, шестичленный андроцей — А6, а в том случае, если их число в цветках одного и того же вида непостоянно (обычно больше 12), — значком ~ или >.
В случае срастания между собой одноименных членов число заключают в скобки (сросшийся пятичленный венчик — Со(5), двубратственный андроцей — А(5+4), 1). Если они расположены несколькими кругами, то цифры, указывающие на число членов в отдельных кругах, соединяют знаком "плюс" (Р3+3).
При обозначении верхней завязи под цифрой числа плодолистиков ставят черточку, при обозначении нижней завязи ее ставят над цифрой. Например, формулы цветков: яблони — *Ca5Co5A~G( 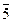 ), гороха — Са(5)Со1,2,2А(5+4),1G1,, лилии — *P3+3A3+3G(3), огурца —
), гороха — Са(5)Со1,2,2А(5+4),1G1,, лилии — *P3+3A3+3G(3), огурца — 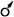 *Са(5)Со(5)А(2),(2),1;
*Са(5)Со(5)А(2),(2),1; 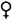 *Са(5)СО(5)G(
*Са(5)СО(5)G( 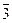 )
)
Диаграммой называют схематическую проекцию цветка на плоскости, при этом цветок мысленно пересекается поперек перпендикулярно его оси. Принят единый способ ориентации диаграммы: ось соцветия вверху, а кроющий лист внизу. На диаграмме части цветка имеют условные обозначения (рис. 133).
Части околоцветника на диаграмме обозначаются дугами: чашелистики — с выступом на середине дуги, лепестки — без выступа. Тычинки обозначаются в форме поперечного разреза пыльника или тычиночной нити; гинецей — в виде поперечного разреза завязи. В случае срастания между собой отдельных членов фигуры, обозначающие их на диаграмме, соединяются дугами.
Соцветия
Соцветие — это система видоизмененных побегов покрытосеменного растения, несущих цветки. Биологическое преимущество соцветий перед одиночными цветками несомненно. Оно заключается в повышении гарантии перекрестного опыления как у энтомофильных, так и анемофильных растений. Любое соцветие имеет главную ось (ось соцветия) и боковые оси, которые или не разветвлены, или в разной степени ветвятся. Конечные их ответвления (цветоножки) несут цветки.
Оси соцветия делятся на узлы и междоузлия. На узлах соцветия располагаются прицветники, а на узлах цветоножки — прицветнички (рис. 134).
При классификации соцветий учитывают наличие и характер листьев, порядок ветвлений осей, способ нарастания осей, поведение апикальных меристем главной и боковых осей.
В зависимости от степени разветвления соцветия делят на простые и сложные. У простых соцветий на главной оси располагаются одиночные цветки и, таким образом, ветвление не превышает двух порядков (гиацинт, черемуха, подорожник, первоцвет, подсолнечник). У сложных соцветий на главной оси расположены не одиночные цветки, а боковые оси частных (парциальных, или элементарных) соцветий, т. е. ветвление достигает трех, четырех и т. д. порядков (морковь, сирень, бирючина, калина, пшеница, мятлик). Нарастание осей может быть моноподиальным или симподиальным. При моноподиалъном нарастании каждая ось формируется за счет деятельности апикальной меристемы, являясь побегом одного порядка. При этом боковые цветки расцветают снизу вверх (акропетально) или центростремительно, если "ось" сильно укорочена и блюдцевидно уплощена. Верхний боковой цветок раскрывается последним. Такие соцветия называют моноподиальными, рацемозными или ботрическими (лат. racemus — кисть, греч. ботрион —кисть). Моноподиальные соцветия имеются, например, у черемухи, пастушьей сумки.
При симподиальном нарастании оси являются составными, представляя собой совокупность побегов нескольких порядков. Первым раскрывается верхушечный цветок на оси первого порядка, вторым — на оси второго порядка и т. д. (базипетально). Иными словами, распускание цветков идет от верхушки к боковым ветвям, или центробежно, если оси расположены в одной плоскости. Такие соцветия — симподиальные, или цимозные (лат. цима — полузонтик), имеются, например, у картофеля, незабудки.
При описательной классификации соцветий, которую используют в справочниках, определителях и пособиях по систематике растений, учитывают два признака: характер (порядок) ветвления и способ нарастания осей.
Простые кистевидные соцветия (ботрические) (рис. 135). Обычно Это моноподиальные соцветия, классификация которых связана с длиной и формой главной оси, а также наличием или отсутствием и Длиной цветоножек.
8.11.1. Простые соцветия с удлиненной осью. Кисть (основной вариант простых соцветий) — ось тонкая с цветками на хорошо выраженных цветоножках более или менее одинаковой длины. Они бывают фроидозные (фиалка трехцветная), брактеозные Слюпин, черемуха), голые (капуста, сурепка), открытые (гиацинт), Закрытые (колокольчик персиколистный).
Щиток — соцветие, сходное с кистью, но у цветков которого Цветоножки разной длины (у нижних цветков они намного длиннее верхних), поэтому все цветки расположены в одной плоскости (груша).
Колос — производное кисти, отличающееся от нее сидячими цветками (подорожник, ятрышник).
Сережка — повислый колос, т. е. колос с мягкой осью, несущий однополые цветки; после цветения соцветие обычно целиком опадает (ива, тополь).
Початок — разновидность колоса, но с толстой мясистой осью (белокрыльник, аир, антуриум). Часто початок окружен листом разной формы и окраски, который называют покрывалом или крылом.
Простые соцветия с укороченной осью. Зонтик — производное кисти с сильно укороченной тонкой осью и цветками на цветоножках одинаковой длины (чистотел, примула, женьшень).
Головка — видоизмененный зонтик с сидячими или почти сидячими (с очень короткими цветоножками) цветками (клевер, люцерна хмелевая). Ось соцветия булавовидно расширена.
Корзинка — близка к головке. Характерна для представителей семейства Астровые. Укороченная ось корзинки разрастается в виде блюдца или конуса, на ней располагаются плотно сомкнутые мелкие сидячие цветки. Такая ось называется ложем соцветия (ее неудачно называют общим цветоложем, но цветоложе — ось цветка). Снизу и с боков ложе соцветия окружено оберткой, составленной сближенными присоцветными листьями верховой формации. Строение листьев оберток очень разнообразно. Они защищают молодые, еще не раскрывшиеся корзинки, часто способствуют распространению плодов, цепляясь за шерсть животных благодаря крючковидной верхушке (лопух). Внешний вид корзинок имитирует облик одиночных цветков: обертка похожа на чашечку, яркие краевые цветки — на венчик. Поскольку корзинка — соцветие моноподиальное, распускание цветков в ней идет центростремительно — от периферии к центру.
Сложные соцветия.
Ботрические. Сложные ботрические соцветия могут быть как открытыми, так и закрытыми.
Сложная кисть — соцветие, у которого на удлиненной моноподиальной главной оси располагаются ботрические парциальные соцветия, являющиеся простыми кистями. В зависимости от степени ветвления, т. е. сложности парциальных соцветий, существуют разные типы сложных кистей Двойная кисть — соцветие, у которого на главной оси располагаются простые кисти (донник, вероника простертая, чемерица зеленая). Тройная кисть — где простые кисти имеют оси не второго, а третьего порядка (вайда красильная, верблюжья колючка, хрен, алоэ крупноплодное).
Сложный колос — соцветие, морфологически близкое к двойной кисти. В нем на удлиненной главной оси располагаются простые колосья, которые называют колосками (пшеница, рожь, ячмень).
Сложный зонтик — другое производное двойной кисти. В нем главная ось укорочена, и на ней расположена розетка верховых листьев, называемая общей оберткой. Из пазух листьев выходят лучи сложного зонтика, завершающиеся парциальными соцветиями — простыми зонтиками, которые называют зонтичками. Цветки в зонтичках часто имеют выраженные прицветники, составляющие частную обертку (оберточку). Сложные зонтики свойственны многим растениям семейства Сельдерейные (морковь, петрушка, укроп, сельдерей). Это открытые соцветия, поэтому цветки в них распускаются центростремительно.
Метелка — отличается обильным ветвлением и тем, что нижние парциальные соцветия ветвятся гораздо сильнее верхних и сильнее развиты, вследствие чего имеет пирамидальную форму (мятлик, сирень, гортензия метельчатая).
Составные (агрегатные) соцветия. Характеризуются тем, что у них тип сложного соцветия отличается от типа частных соцветий. Например, метелка корзинок (полынь обыкновенная), кисть зонтиков (плющ обыкновенный), колос корзинок (сушеница лесная).
Цимозные соцветия. Цимоиды — закрытые соцветия, у которых главная ось не выражена, а парциальные соцветия развиваются только в непосредственной близости от верхушечного цветка, а остальные редуцированы (рис. 136). В цимоидах первыми раскрываются цветки, заканчивающие оси более низких порядков. Цимоиды разделяют в зависимости от числа боковых осей, сменяющих в ходе симподиального нарастания одну материнскую на три основных типа (монохазий, дихазий, плейохазий).
Монохазий (однолучевой верхоцветник) — такой цимоид, у которого каждая материнская ось несет одну дочернюю; в простейшем случае под цветком, завершающим ось предыдущего порядка, формируется один цветок следующего порядка. В зависимости от направления осей различают два типа монохазиев: завиток и извилина. Завиток (улитка) — все оси направлены в одну сторону (незабудка, красоднев); завиток с сильно укороченными осями называют клубком (грыжник голый).
Извилина — оси более высоких порядков возникают попеременно то в одну, то в другую сторону по отношению к осям более низких порядков (лаперузия, норичник, петуния).
Дихазий (двухлучевой верхоцветник) — соцветие, у которого каждая ось предыдущего порядка несет две оси следующего порядка, т. е. под верхушечным цветком главной оси развиваются два парциальных соцветия (звездчатка, ясколка).
Плейохазий (многолучевой верхоцветник) — соцветие, у которого каждую ось предыдущего порядка сменяют более двух осей следующего порядка (родиола).
Циатий — особый тип цимозного соцветия с раздельнополыми цветками (молочайные). Циатий состоит из верхушечного пестичного цветка (редуцированного пестичного соцветия) и пяти тычинок (редуцированных пяти тычиночных парциальных соцветий). Он окружен оберткой, состоящей из кроющих листьев парциальных соцветий.
В сложных соцветиях нередко сочетаются разные способы нарастания осей.
Тирсы — сложные соцветия с моноподиально нарастающей главной осью и симподиально нарастающими боковыми соцветиями (цимоидами). Степень разветвления боковых соцветий уменьшается от основания к верхушке, придавая тирсу пирамидальную форму.
Опыление
Опыление — это перенос пыльцы с тычинок на рыльце пестика. Простейший и, вероятно, древнейший способ опыления у растений только с обоеполыми цветками — самоопыление, или автогамия (греч. авто — сам), при котором пыльца опыляет рыльце пестика того же цветка. При перекрестном опылении (аллогамия; греч. аллос — другой) пыльца опыляет рыльца других цветков. Различают две формы перекрестного опыления: гейтоногамия (греч. гейтон—сосед), когда оно происходит в пределах одного растения, но пыльца с одного цветка попадает на рыльце пестика другого; ксеногамия (греч. ксенос — чужой), или собственно перекрестное, когда пыльца с цветка одной особи переносится на рыльце пестика другой особи. С генетической точки зрения гейтоногамия равноценна автогамии, так как при этом варианте перекрестного опыления не происходит рекомбинации индивидуальных признаков.
8.12.1. Самоопыление.Играет важную роль в стабилизации признаков вида как средство размножения и расселения генетически однородных популяций, в селекции — при выведении чистых линий. Самоопылителями являются очень многие виды культурных растений: пшеница, ячмень, овес, просо, горох, фасоль, бобы, хлопчатник, лен, томат и др.
Самоопыление происходит как у раскрывшихся — хазмогамных (греч. хазма — трещина, зев) цветков (сельдерейные), так и у закрытых — клейстогамных (греч. клейстос — закрытый) цветков (арахис, фиалка, истод, кислица).
Однако биологические преимущества ксеногамии очевидны, так как при этом возрастают возможности рекомбинаций генетического материала и это способствует увеличению внутривидового разнообразия и дальнейшей приспособительной эволюции.
Для эволюционного процесса оптимально сочетание самоопыления и перекрестного опыления. Однако преобладает собственно перекрестное, свойственное 90 % растений.
У ряда цветковых растений есть специальные приспособления морфологического и физиологического характера, предотвращающие или ограничивающие самоопыление. Рассмотрим некоторые из них.
Двудомность — наиболее надежное средство, препятствующее самоопылению. Однодомность устраняет автогамию, но не предохраняет от генетически равноценной ей гейтоногамии. Дихогамия (греч. дихе — на две части) — разновременное созревание пыльцы и рыльца в одном цветке. Дихогамия встречается в двух формах: протерандрия и протерогиния. Протерандрия — более раннее вскрывание пыльников, когда рыльце еще не созрело. Протерогиния — более раннее созревание рыльца, т. е. рыльце становится восприимчивым к пыльце, когда пыльники еще не вскрылись. Протерандрия встречается чаще (бобовые, астровые, гвоздичные, сельдерейные, мятликовые, осоковые); протерогиничны цветки у норичниковых, гречихи, белены, гладиолуса, некоторых мятликовых и осок.
Гетеростилия (разностолбчатость) состоит в том, что у некоторых видов (первоцвет, дикорастущие виды гречихи, льна, некоторые бурачниковые, маревые) имеется два типа внешне похожих цветков, но различающихся по длине столбиков и тычиночных нитей. У одних особей столбики пестиков длинные и пыльники располагаются низко в трубке венчика, у других — соотношение обратное (рис. 137).
Вероятно, при посещении особей с морфологически разными цветками пыльца высыпается на разные участки тела насекомого и соответственно может попасть затем только на соответствующее по высоте рыльце, т. е. из короткостолбчатого цветка в длинностолбчатый, и наоборот. Таким образом, гетеростилия служит защитой от гейтоногамии.
Самонесовместимость (самостерильность или физиологическая несовместимость) выражается в подавлении прорастания пыльцы на рыльце пестика той же особи при автогамии и гейтоногамии (некоторые сорта яблони и др.). В практике плодоводства для каждого такого сорта экспериментально определены сорта-опылители. Поэтому при закладке плодового сада рядом с самостерильными сортами сажают сорта-опылители. У насекомоопыляемых растений с обоеполыми цветками Морфологическим препятствием к самоопылению служит расположение пыльников и рылец, исключающее попадание пыльцы на рыльце. Так, например, в цветках норичниковых, яснотковых, астровых, колокольчиков, горечавок, лилий столбики далеко высовываются за пределы околоцветника, а тычинки находятся глубоко внутри него.
С другой стороны, часто осуществляется принцип "лучше самоопыление, чем никакого опыления", поэтому в цветках астровых, колокольчиков, горечавок и др. в случае отсутствия перекрестного опыления лопасти рылец, разрастаясь, загибаются вниз и соприкасаются с пыльцой.
8.12.2. Перекрестное опыление.Механизмы перекрестного опыления разнообразны. Их подразделяют на два основных типа: абиотическое — с помощью неживых факторов внешней среды и биотическое — с помощью животных. Абиотическое опыление связано прежде всего с переносом пыльцы ветром (анемофилия),а также водой (гидрофилия). Анемофилы — это главным образом растения открытых пространств. У них, как правило, цветки мелкие с невзрачным или редуцированным околоцветником; тычинки и рыльца пестиков выступают за его пределы; часто рыльца мохнатые, а их воспринимающая поверхность сильно увеличена. Пыльца очень легкая, благодаря чему слабыми токами воздуха она разносится на расстояние до 5 км и более. Пыльцы образуется очень много (например, одно растение кукурузы формирует 50 000 000 пылинок). У некоторых ветроопыляемых растений, например у ржи, формируются качающиеся пыльники, которые в период цветения свисают на тонких длинных тычиночных нитях и рассеивают пыльцу. У анемофилов очень часто цветки собраны в многоцветковые соцветия.
Деревья и кустарники цветут до того, как развернутся листья.
Гидрофилия свойственна главным образом водным растениям с цветками, целиком погруженными в воду (роголистники, наяда, морская трава). У них пыльники часто нитевидные или червеобразные, что облегчает передвижение в воде и опыление; они лишены экзины, предохраняющей от высыхания.
В зависимости от агентов различают следующие виды биотического опыления: перенос насекомыми — энтомофилия, птицами — орнитофилия, летучими мышами — хироптерофилия и др.
Энтомофилия. Насекомыми опыляется около 70 % всех покрытосеменных растений. Они сыграли важнейшую роль в эволюции цветка, которая шла по пути тесного приспособления цветка и насекомого друг к другу. Иногда связь становится столь тесной, что растение оказывается в полной зависимости от опылителя и не в состоянии произрастать при отсутствии опылителя.
Орнитофилия. В тропиках цветки многих растений опыляют мелкие птицы (колибри, медососы), питающиеся нектаром. Цветки таких растений обычно крупные, ярко-красной окраски, которую лучше различают птицы; вырабатывают значительное количество водянистого нектара.
Оплодотворение
Между опылением и оплодотворением (слиянием гамет) у разных растений проходит определенное время: у большинства растений — до двух суток, у кок-сагыза — 15...45 мин, у орешника — три-четыре месяца. Оплодотворению предшествует прорастание пылинки, которое начинается с выхода из апертуры пыльцевой трубки. По мере ее роста по столбику пестика в ее растущий конец переходят ядро сифоногенной клетки и два спермия (рис. 138). Достигнув завязи, пыльцевая трубка направляется к семязачатку и проникает в него чаще всего через микропиле.
После проникновения в зародышевый мешок пыльцевой трубки оболочка на ее кончике разрывается и ее содержимое изливается внутрь. При этом один из спермиев сливается с яйцеклеткой, образуя диплоидную зиготу, а второй— с центральным (вторичным) ядром зародышевого мешка, образуя триплоидное ядро, из которого формируется запасающая ткань — эндосперм. Так происходит двойное оплодотворение, характерное только для покрытосеменных. Прочие клетки зародышевого мешка (синергиды и антиподы) дегенерируют.
Двойное оплодотворение было открыто русским ботаником С. Г. Навашиным в 1898 г. при исследовании эмбриогенеза у лилии лесной и было оценено как одно из крупнейших открытий в области естественных наук XIX в. Биологическое значение двойного оплодотворения очень велико.
В отличие от голосеменных, у которых довольно мощный гаплоидный эндосперм развивается до оплодотворения, у покрытосеменных триплоидный эндосперм формируется только после оплодотворения. Этим достигается существенная экономия энергетических ресурсов организма. Семязачатки покрытосеменных, не обремененные запасающей питательной тканью впрок, развиваются гораздо быстрее, чем у голосеменных.
Вопросы для самоконтроля
1. Что такое цветок? Из каких частей он состоит? Каковы их функции?
2. Что такое опыление? Какие типы опыления известны?
3. Какие приспособления препятствуют самоопылению?
4. Какие бывают группы соцветий? В чем их принципиальное отличие?
5. Что такое микроспорогенез и мегаспорогенез? Что является гомологом мегаспорангия?
6. Что такое микрогаметогенез и мегагаметогенез?
7. Что является гомологом мужского гаметофита?
8. Чему гомологичен зародышевый мешок?
Резюме.
Существует два типа размножения: половое и бесполое. При половом размножении особи нового поколения появляются при участии двух физиологически различных организмов. . Процесс слияния гамет называется оплодотворением. Растение, на котором образуются гаметы, —гаметофит. Процесс формирования гамет — гаметогенез — происходит в особых органах — гаметангиях. Гаметофиты равноспоровых растений обоеполые, несут мужские и женские гаметангии. У разноспоровых растений из микроспор развиваются гаметофиты с мужскими гаметангиями, а из мегаспор — с женскими. В жизненном цикле каждого растения, имеющего половое размножение, существует смена ядерных фаз — гаплоидной и диплоидной. Переход от гаплоидного состояния к диплоидному происходит в результате полового процесса при образовании зиготы; от диплоидного к гаплоидному — в результате мейоза обычно при спорообразовании. Оплодотворение и мейоз взаимосвязаны, это две стороны жизненного процесса, поддерживающие постоянство числа хромосом. У высших растений и ряда водорослей происходит чередование поколений — бесполого (спорофита) и полового (гаметофита). На диплоидном спорофите за счет мейотического деления образуются гаплоидные споры. Из споры развивается гаплоидный гаметофит, производящий гаплоидные гаметы. При их слиянии в зиготе восстанавливается диплоидный набор хромосом. Из зиготы вновь развивается диплоидный спорофит.
Собственно бесполое размножение происходит при помощи специализированных клеток — спор, при прорастании которых развиваются новые особи. Вегетативное размножение - увеличение числа особей за счет отделения жизнеспособных частей вегетативного тела и их последующей регенерации (восстановления до целого организма).
Цветок — это сложный репродуктивный орган покрытосеменных (цветковых) растений. Цветок имеет ось, или цветоложе, несущее листочки околоцветника, тычинки и пестик или пестики. Пестик состоит из одного или нескольких плодолистиков. Главные части пестика — замкнутая завязь с семязачатками внутри и воспринимающее пыльцу рыльце. Принципиальное отличие цветка от шишки голосеменных состоит в том, что семязачатки находятся внутри завязи и пыльца попадает при опылении на рыльце, а не непосредственно на семязачаток. Двойной околоцветник дифференцирован на чашечку и венчик. Простой околоцветник не дифференцирован на чашечку и венчик, состоит из совокупности однородных листков околоцветника. Если простой околоцветник состоит из зеленых листков, то он называется чашечковидным (крапива, конопля); если из иначе окрашенных — венчиковидным (гречиха, лук, тюльпан).
Андроцей — это совокупность тычинок (микроспорофиллов) одного цветка. Тычинка состоит из тычиночной нити, посредством которой она нижним концом прикреплена к цветоложу, и пыльника на ее верхнем конце.
Процесс образования микроспор в микроспорангиях, которыми являются гнезда пыльника, называется микроспорогенезом. Процесс образования из микроспоры мужского гаметофита (пылинки) называется микрогаметогенезом.
Гинецей — это совокупность плодолистиков или карпелл в цветке, образующих один или несколько пестиков. Пестик — наиболее существенная часть цветка, из которой формируется плод, возникает из плодолистика или плодолистиков вследствие смыкания и срастания их краев. Завязь - это замкнутая нижняя расширенная полая, наиболее важная часть пестика, несущая семязачатки. Столбик - тонкая цилиндрическая стерильная часть пестика, отходящая обычно от верхушки завязи. Рыльце - расширенная часть на верхушке столбика, предназначенная для восприятия пыльцы.
Гинецей, состоящий из одного плодолистика, называют монокарпным, состоящий из нескольких свободных (несросшихся) плодолистиков, каждый из которых образует свой пестик, называют апокарпным, образованный при срастании нескольких плодолистиков, называют ценокарпным. В зависимости от способа срастания плодолистиков различают несколько ценокарпных гинецеев: синкарпный, лизикарпный и паракарпный.
Семязачатки — это небольшие образования, которые располагаются в завязи пестика. В семязачатке происходят: мегаспорогенез — формирование мегаспор; мегагаметогенез — формирование женского гаметофита и процесс оплодотворения. После оплодотворения (реже без него) семязачаток развивается в семя. В зависимости от ориентации, взаимного расположения частей и степени изогнутости нуцеллуса различают пять основных типов семязачатка: ортотропный (прямой) — микропиле и фуникулюс расположены на противоположных концах оси семязачатка; анатропный (обратный или обращенный) — нуцеллус повернут по отношению к прямой оси семязачатка на 180°, вследствие чего микропиле и фуникулюс расположены рядом; гемитропный (полуповернутый) — семязачаток повернут на 90°, вследствие чего микропиле и нуцеллус расположены по отношению к фуникулюсу под углом 90°; кампилотропный (односторонне изогнутый) — нуцеллус изогнут только односторонне микропилярным концом, поэтому микропиле и фуникулюс расположены рядом; амфитропный (двусторонне изогнутый) — нуцеллус двусторонне изогнут в виде подковы, при этом микропиле и фуникулюс расположены тоже рядом.
Мегаспоры образуются в процессе мегаспорогенеза в нуцеллусе (мегаспорангий) семязачатка. После этого начинается мегагаметогенез, т. е. формирование женского гаметофита — зародышевого мешка.
Сущность цветения состоит во вскрывании пыльников и функционировании рылец пестиков как органов, воспринимающих пыльцу.
Строение цветка можно выразить в виде формулы и диаграммы. Диаграммой называют схематическую проекцию цветка на плоскости, при этом цветок мысленно пересекается поперек перпендикулярно его оси. Принят единый способ ориентации диаграммы: ось соцветия вверху, а кроющий лист внизу.
Соцветие — это система видоизмененных побегов покрытосеменного растения, несущих цветки. При классификации соцветий учитывают наличие и характер листьев, порядок ветвлений осей, способ нарастания осей, поведение апикальных меристем главной и боковых осей. В зависимости от степени разветвления соцветия делят на простые и сложные.
Опыление — это перенос пыльцы с тычинок на рыльце пестика. Самоопыление играет важную роль в стабилизации признаков вида как средство размножения и расселения генетически однородных популяций, в селекции — при выведении чистых линий. Перекрестного опыления подразделяют на два основных типа: абиотическое — с помощью неживых факторов внешней среды и биотическое — с помощью животных.
Двойное оплодотворение было открыто русским ботаником С. Г. Навашиным в 1898 г. при исследовании эмбриогенеза у лилии лесной и было оценено как одно из крупнейших открытий в области естественных наук XIX в. После проникновения в зародышевый мешок пыльцевой трубки оболочка на ее кончике разрывается и ее содержимое изливается внутрь. При этом один из спермиев сливается с яйцеклеткой, образуя диплоидную зиготу, а второй— с центральным (вторичным) ядром зародышевого мешка, образуя триплоидное ядро, из которого формируется запасающая ткань — эндосперм. Так происходит двойное оплодотворение, характерное только для покрытосеменных. Прочие клетки зародышевого мешка (синергиды и антиподы) дегенерируют. У покрытосеменных триплоидный эндосперм формируется только после оплодотворения.
Дата добавления: 2020-10-14; просмотров: 1031;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
