Взаимодействие фага с бактериальной клеткой.
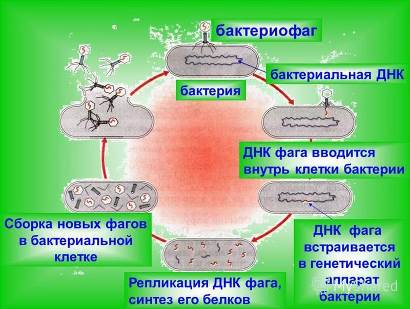
При контакте бактерии со свободным фагом заражение произойдет, если бактериальная клетка будет чувствительна к вирусу. Вначале фаг адсорбируется на оболочке клетки бактерии и оболочка растворяется при помощи фермента лизоцима. Затем благодаря сокращению хвостового отростка нуклеиновая кислота фага впрыскивается через хвостовой отросток в цитоплазму клетки. Фаги остаются адсорбированными на оболочке бактерии, но их головки выглядят опустевшими.
Несколько минут после проникновения нуклеиновой кислоты внутрь клетки бактерии (скрытый период) фаговые частицы не обнаруживаются. Однако в клетке развиваются процессы, обусловленные фаговым геномом. В этот период начинается синтез новых (ранних) белков. К ним относятся ферменты, необходимые для синтеза ДНК фага. Синтезируется несколько сотен новых фаговых ДНК. Новые молекулы ДНК фага образуются отчасти из материала питательной среды и отчасти из деградированной ДНК хозяина. На поздней стадии латентного периода образуются «поздние» белки, которые включают субъединицы фаговой головки и хвостовой части, а также лизоцим.
Синтез всех ферментов и фаговых структурных белков кодируется фаговой ДНК. В конце латентного периода белковые субъединицы головки и хвостовой части фага самопроизвольно агрегируются (самосборка) с образованием полного капсида. На последнем этапе размножения происходит формирование зрелых частиц фага, которое состоит в необратимом объединении фаговой ДНК и белковой оболочки. После этого бактериальные клетки лизируются и фаги освобождаются в окружающую среду. Зрелые частицы представляют собой морфологически типичные бактериофаги, которые могут заражать новые клетки. В каждой зараженной клетке весь цикл репродукции повторяется снова от начала до конца.
По характеру взаимодействия с клеткой бактерии бактериофаги делятся на вирулентные и умеренные. Вирулентные фаги всегда лизируют клетку бактерии.
Умеренные фаги могут вызвать лизис клетки бактерии, но могут перейти и в неинфекционную форму. В этом случае молекула ДНК фага прикрепляется к ДНК бактерии путем кроссинговера и передается с нею дочерним клеткам. Фаг, существующий в такой форме, называется профагом.
Клетки бактерий, имеющие в своей хромосоме профаг, называются лизогенными, а явление совместного существования ДНК бактерии и профага называется лизогенией.
Профаг может сосуществовать с бактериальной клеткой длительное время, но при определенных условиях он может отделиться от ДНК бактерии, перейти в вирулентную форму и вызвать лизис бактериальной клетки. РНК-вирусы, так же как и ДНК-вирусы, могут вызвать лизогенное состояние клеток бактерий. На РНК вируса может синтезироваться комплементарная ей ДНК, на которой синтезируется вторая цепь ДНК. Таким образом образуется полноценная молекула ДНК, способная соединиться с ДНК клетки хозяина. В качестве провируса эта ДНК может передаваться потомству, и вызываемое данным вирусом заболевание может стать как бы наследственным. Наличие профага в составе бактериальной хромосомы не мешает репликации ДНК бактериальной клетки. Однако гены профага, встроенные в ДНК клетки, не транскрибируются. Это связано с образованием в клетке бактерии репрессора – низкомолекулярного белка, блокирующего считывание наследственной информации, записанной в фаговой ДНК. Синтез репрессора контролируется генами профага. Умеренные фаги могут быть дефектными, т. е. неспособными к образованию зрелых фаговых частиц. Такие фаги осуществляют трансдукцию и используются в генной инженерии.
Дата добавления: 2020-10-01; просмотров: 717;
Поиск по сайту
Узнать еще
- А.2 Взаимодействие движителя с грунтом.
- В отличие от координации, взаимодействие исполняется на уровне отдельных предприятий.
- Взаимодействие (перенос) навыков.
- Взаимодействие аллельных генов
- Взаимодействие анализаторов при приеме информации
- Взаимодействие бактериофага с бактериальными клетками
- Взаимодействие банков и страховых компаний
- Взаимодействие биотических и абиотических факторов. Выживание и воспроизводство. Адаптации, их виды. Принцип коэволюции. Монокультура и природа.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
