Понятие о генотипе и фенотипе микроорганизмов.
Совокупность генов бактериальной клетки называется ее генотипом. Совокупность всех признаков и свойств, проявляемых данной культурой, называется ее фенотипом. В отличие от особей высших организмов, у которых исследуются признаки и свойства каждой особи, у микроорганизмов изучаются признаки и свойства в целом всей культуры, т. е. совокупности клеток, включающих миллионы и миллиарды особей.
Культуры микробов могут отличаться морфологическими, физиологическими и биохимическими признаками. К морфологическим признакам относятся окраска, размер, форма, характер края и поверхности отдельно растущих колоний и т. д. К физиологическим признакам относится способность или неспособность расти при пониженной или повышенной температуре, устойчивость к антибиотикам, к различным ядам, к облучению и т. д. Биохимическим признаком является отношение микроорганизмов к питательным средам. По способу питания бактерии делятся на прототрофные и ауксотрофные. Прототрофные бактерии могут жить на минимальной питательной среде (содержащей минеральные соли и углеводы) и необходимые им вещества способны синтезировать сами. Ауксотрофные – лишены способности синтезировать одну или несколько аминокислот или другие факторы роста из более простых предшественников. Такие штаммы можно идентифицировать по его неспособности расти и размножаться на синтетической среде, не содержащей какого-то специфического фактора роста. Таким образом, культура микроорганизма может быть изучена в отношении многих признаков.
Гены, ответственные за синтез определенного соединения, обозначают тремя строчными буквами, соответствующими начальным буквам этого соединения. Гены исходного дикого типа обозначают со знаком плюс, например his+ - гистидиновый ген, leu+ — лейциновый ген и т. д. Гены чувствительности или резистентности к лекарственным препаратам, фагам и ядам обозначают буквами s (sensitive – чувствительный) и r (resistant— резистентный). Например, чувствительность к стрептомицину обозначают strs, а резистентность strr. Фенотип бактерий обозначают теми же символами, что и генотип, но с прописной буквы. Так, генотипу his+, strr соответствует фенотип Нis+, strr.
Генотип микроорганизмов представлен совокупностью генов, обусловливающих потенциальную возможность формирования любого их признака. Но формирование признака происходит в определенных условиях окружающей среды, которые не всегда способствуют проявлению генотипа. Так, два штамма Е. соlі с генотипами 1ас+ и 1ас- на среде с лактозой будут иметь разные фенотипы: бактерии с генотипом 1ас+ образуют колонии красного цвета, а бактерии с генотипом 1ас- - бесцветные колонии, так как они не ферментируют лактозу. При выращивании этих штаммов на среде без лактозы фенотип их будет одинаковый.
Микроорганизмам присуща большая изменчивость. Изменения, возникающие под влиянием окружающей среды и не сохраняющиеся при переносе клеток в исходные условия, носят название модификационных. Модификации ненаследственны, они не затрагивают генотипа микроба и исчезают в первом или последующих поколениях. Микроорганизмам, как и всем прочим живым организмам, свойственно проявление комбинативной изменчивости, которая является наследственной. В силу того что прокариоты имеют гаплоидный набор хромосом и им несвойственно половое размножение, генетические рекомендации у них имеют свои особенности.
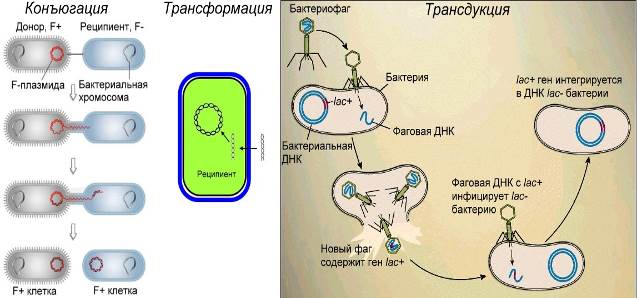
Рекомбинация у бактерий происходит путем переноса генетического материала из клетки донора в клетку реципиента при помощи трансформации, трансдукции и конъюгации. Эти процессы отличаются друг от друга главным образом механизмом переноса генетического материала.
Трансформация
В процессе трансформации принимают участие две бактериальные клетки – донор и реципиент. Трансформирующий агент представляет собой часть молекулы ДНК донора, которая внедряется в геном реципиента, изменяя его фенотип. В процессе трансформации клетки донора и реципиента не соприкасаются друг с другом. Механизм переноса генетического материала заключается в том, что из клеток донора выделяются в окружающую среду молекулы или фрагменты молекул ДНК. Сначала ДНК адсорбируется на оболочке клетки реципиента. Затем через определенные рецепторные участки ее стенки при помощи специальных клеточных белков ДНК втягивается внутрь клетки. Проникающая донорская ДНК должна быть двухцепочной. В реципиентной клетке она становится одноцепочной. В ДНК реципиента включается одна из цепей трансформирующего фрагмента. Эта цепь вступает в синапсис с гомологичным участком хромосомы реципиента и встраивается в нее посредством кроссинговера. При этом участок ДНК реципиента замещается фрагментом донора. Молекула ДНК со вставкой трансформирующего участка оказывается гибридной. При следующем удвоении возникает одна нормальная дочерняя молекула ДНК, другая трансформированная.
Способность бактерий-реципиентов к трансформации определяется их физиологическим состоянием. Такое физиологическое состояние было названо компетентностью. Состояние компетентности краткосрочно и приурочено к определенному времени клеточного цикла. Трансформирующей способностью обладают только крупные молекулы ДНК с молекулярной массой 106 Д и более. Кроме того, необходимо, чтобы ДНК донора была полностью или частично гомологична ДНК реципиента.
У бактерий трансформация имеет место в пределах одного вида, но наблюдается и между разными близкими видами. Это указывает на то, что у них сохранилась гомологичность некоторых участков ДНК.
Трансдукция
Перенос генов из одной бактериальной клетки в другую при помощи умеренных фагов называется трансдукцией.
Трансдудирующие фаги известны у многих бактерий. Как правило, трансдуцируется один ген, реже два сцепленных гена и очень редко три. При переносе генетического материала происходит замена участка молекулы ДНК фага. Фаг при этом теряет свой собственный фрагмент и становится дефектным. Включение генетического материала в хромосому бактерии реципиента осуществляется механизмом типа кроссинговера. Происходит обмен наследственным материалом между гомологичными участками хромосомы реципиента и материала, привнесенного фагом. Различают три вида трансдукции: общую, (неспецифическую), специфическую и абортивную. При неспецифической трансдукции в период сборки фаговых частиц в их головку вместе с фаговой ДНК может включиться любой из фрагментов ДНК пораженной бактерии. В результате в реципиентные клетки могут переноситься различные гены бактерии донора. При специфической трансдукции профаг включается в определенное место хромосомы бактерии и трансдуцирует определенные гены, расположенные в хромосоме клетки донора рядом с профагом. Частота общей трансдукции составляет от 1 на 1 млн. до 1 на 100 млн. Специфическая трансдукция происходит чаще. Установлено, что фрагмент хромосомы донора, перенесенный в клетку реципиента, не всегда включается в хромосому реципиента, а может сохраняться в цитоплазме клетки, хотя и не размножается. При делении бактерий он попадает только в одну из дочерних клеток. Такое состояние получило название абортивной трансдукции.
Конъюгация
Конъюгацией называется передача генетического материала от одних бактерий другим при их скрещивании. Установлена неравноценная роль родительских штаммов при конъюгации. Выяснилось, что один штамм является донором (мужским), а другой — реципиентом (женским). Клетки-доноры обладают половым фактором F. Он является конъюгативной плазмидой и представляет собой циркулярно замкнутую молекулу ДНК. Половой фактор F автономно существует в цитоплазме. Бактериальные клетки с фактором F обозначаются F +, не имеющие его — F-. Они неспособны быть донорами и являются реципиентами генетического материала.
При конъюгации фактор F может перейти из мужской в женскую клетку и превратить ее в F + без изменения других свойств клетки бактерии. Половой фактор F обладает способностью включаться в геном бактерии. Для этого кольца ДНК генома и плазмиды разрываются в определенной точке и ДНК плазмиды (фактора F) петлеобразно встраивается путем кроссинговера. Фактор Р+ из цитоплазматической структуры превращается в фрагмент хромосомы.
При конъюгации клетки-доноры F+ соединяются с клетками-реципиентами F- при помощи конъюгационного мостика – особой протоплазматической трубки, образуемой клеткой F +. В клетке донора под влиянием фермента эндонуклеазы в точке внедрения фактора F происходит разрыв цепи ДНК. Свободный конец одной из цепей ДНК постепенно начинает передвигаться через конъюгационный мостик в клетку реципиента (F-) и сразу же достраивается до двухцепочной структуры. На оставшейся в клетке-доноре цепи ДНК синтезируется вторая цепь. Конъюгационный мостик очень хрупкий, легко разрывается и, как правило, вся цепь не успевает перейти. Поэтому и фактор F обычно не передается, так как располагается в конце хромосомы. С более высокой частотой передаются гены, расположенные около начальной О-точки хромосомы донора. Затем ДНК донора в гомологичных участках вступает в контакт с ДНК реципиента и в результате кроссинговера некоторые участки одной цепи ДНК реципиента заменяются фрагментами ДНК донора.
Иногда включенный в хромосому половой фактор F освобождается и при этом может (подобно профагу) захватить с собой прилегающий к нему участок ДНК бактерии.
При конъюгации половой фактор вместе с фрагментом ДНК иногда переходит в женскую клетку, превращая ее в мужскую и передавая ей свойства, контролируемые фрагментом хромосомы донора. Процесс переноса генетической информации при помощи полового фактора называется сексдукцией.
Плазмиды. Кроме половых факторов, которые участвуют в скрещивании, большинство бактерий содержит также другие хромосомные элементы, называемые плазмидами. Они представляют собой кольцевые молекулы ДНК, обладают свойствами репликона: могут реплицироваться с помощью ферментов клетки бактерии независимо от основной хромосомы.
У некоторых непатогенных и патогенных видов бактерий обнаружены факторы резистентности к лекарственным веществам — R-факторы. Их присутствие в цитоплазме бактерий и способность передаваться из поколения в поколение, обусловливают устойчивость бактерий к действию одного или нескольких лекарственных веществ (стрептомицина, тетрациклина и др.). К другому типу плазмид относятся колициногенные факторы (колициногены), которые включают гены, обусловливающие синтез бактерией особых белковых веществ – колицинов.
Плазмиды по конъюгационному мостику могут проникать в другую бактерию, даже другого вида, обусловливая или невосприимчивость к определенным лекарственным веществам, или способность к синтезу колицинов. Это свойство плазмид широко используется в генной инженерии.
Дата добавления: 2020-10-01; просмотров: 781;
Поиск по сайту
Узнать еще
- B.I. Понятие культуры
- I. Понятие о методах воспитания.
- I. Понятие о принципах обучения, их взаимосвязь.
- II. Общее понятие о процессе познания и процессе обучения.
- III.IX. Понятие о стратиграфических и петрографических горизонтах
- IV. Понятие “стохастические и нестохастические” эффекты облучения
- IV.4. Понятие устойчивого развития
- А) Понятие о комплексных числах

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
