Геохимическая роль микроорганизмов
Известно, что оптимальное функционирование любой экосистемы во многом зависит от биоразнообразия живых организмов, составляющих ее трофические звенья. Огромную роль на всех трофических уровнях принадлежит микроорганизмам, и в частности бактериям. Они могут быть участниками, как процессов синтеза нового органического вещества, так и являются редуцентами, ответственными за процессы мацерации и деструкции органических тканей до простых минеральных соединений.
Главный парадокс жизни, по мнению М.М. Камшилова (1974 Эволюция биосферы. Изд. Наука,1974.С. 254) заключается в том, что ее непрерывность обеспечивается процессами распада, деструкцией. Разрушаются сложные органические соединения, освобождается энергия, теряется запас информации, свойственный сложно организованным живым телам. В результате деятельности деструкторов, любая форма жизни неизбежно будет включаться в биотический круговорот. Поэтому с их помощью осуществляется естественная саморегуляция биосферы.
Два свойства позволяют микроорганизмам играть столь важную роль:
-- возможность сравнительно быстро приспосабливаться к различным условиям;
-- способность использовать в качестве источника углерода и энергии самые различные субстраты.
Высшие организмы не обладают такими способностями. Поэтому они могут существовать лишь в качестве своеобразной надстройки на прочном фундаменте одноклеточных.
Конечное деструктивное звено – минерализация органических веществ с возвратом СО2 в атмосферу осуществляется в почвенном покрове Земли за счет деятельности почвенных микроорганизмов. При минерализации органики в атмосферу кроме СО2 возвращаются еще СН4 – метан, водород – Н2 , угарный газ – СО, Н2S – сернистый газ, N2O – закись азота, N2 – азот. Таким образом, роль почвенных микроорганизмов проявляется не только в деструкции органики, но также в контроле газового состава атмосферы, климата и преобразовании литосферы.
Биоэкологический мониторинг подразумевает использование биологических объектов, и бактерий в том числе, для индикации состояния экосистем. Так, например, известно, что определенный уровень биоразнообразия бактериопланктона может отражать степень трофности водоема, а индикаторные виды бактерий могут отражать и санитарное состояние водоема. Доминирующие виды бактерий, обнаруживаемые в воздухе, могут быть индикаторами территорий риска с повышенными концентрациями различных химических соединений, в том числе и токсичных для человека.
Сложившиеся в естественных условиях микробные ценозы и ассоциации утилизируют те или иные естественные природные субстраты в адекватной химической среде. Любое изменение тех или иных элементов среды приводит к интенсификации или угнетению процессов жизнедеятельности микроорганизмов. Вносимые человеком в почву высокие концентрации различных веществ не только губительны для значительной части популяций микробов, но и обладают селективным эффектом, в результате чего в среде начинают преобладать резистентные к токсикантам штаммы, а естественный ход геохимических процессов нарушается.
В общем курсе микробиологии студенты знакомятся с различными аспектами деятельности микроорганизмов. Изучаются процессы утилизации белковой и небелковой органики, физиологическая роль микробов в жизни макроорганизма. Достаточно полно обучающиеся знакомятся с методами взятия проб. К сожалению, большее внимание на лабораторных занятиях приходится уделять задачам санитарно-гигиенического плана. Вместе с тем, практика показывает, что сегодня необходимы знания и навыки исследования микроорганизмов в природных биосистемах.
Предпринятые на кафедре попытки участия студентов в проведении микробиологического мониторинга озер Пустынского заказника оказались вполне успешными и позволили наглядно и увлекательно пробудить интерес студенческой аудитории к подобным исследованиям. Сегодня наряду с традиционными медико-гигиеническими проблемами, решаемыми студентами в процессе выполнения курсовых и дипломных работ появились исследования экологической тематики. Сегодня, на наш взгляд, является целесообразным формировать интерес к подобным задачам уже в рамках общего курса микробиологии, организовав цикл занятий в период летней учебной практики студентов 2 курса.
Из большого числа задач, представляющих образовательный и научный интерес, нами выделены несколько.
1. Определение общего микробного числа (ОМЧ) - один из наиболее доступных и достаточно репрезентативных показателей динамики количества и качества микрофлоры.
2. Определение числа, разнообразия и активности штаммов аммонификаторов позволяют индицировать напряженность процессов отмирания и разложения белковой органики.
3. Выявление активности серобактерий и железобактерий - уникальных фототрофных бактерий, осуществляющих оксигенный и аноксигенный фотосинтез, позволяет судить о напряженности процессов синтеза и деструкции в естественных микробоценозах и экосистемах в целом.
УЧАСТИЕ ПОЧВЕННЫХ МИКРООРГАНИЗМОВ В ЦИКЛАХ ОСНОВНЫХ БИОГЕННЫХ ЭЛЕМЕНТОВ В БИОСФЕРЕ И ПОЧВООБРАЗОВАТЕЛЬНЫХ ПРОЦЕССАХ
Жизнь на нашей планете возникла, развивается и процветает благодаря энергии Солнца. Приблизительно 10% энергии, достигающей поверхности Земли, тратится на испарение воды, что вызывает абиотический, или большой круговорот (БГК). Только 0,1% энергии Солнца закрепляется в синтезируемых растениями органических соединениях, что составляет основу биотического, или малого биологического, круговорота (МБК, см. рис. 1).Солнечная энергия, поглощенная растениями, совершает огромную работу: она «запускает» процессы биосинтеза и трансформируется в энергию химических связей образующихся органических веществ. Главный запас потенциальной биогенной энергии сосредоточен в почвенном покрове Земли в виде корней растений, биомассы микроорганизмов и гумуса.
В биосинтетические процессы вовлекаются разнообразные элементы, которые подвергаются постоянным превращениям. Так как количество этих элементов на нашей планете может быть определено конечными величинами, то «бесконечность» жизни обеспечивается непрерывно идущим круговоротом этих элементов. Различные этапы круговорота элементов осуществляются разными группами организмов.
Фотосинтетическое связывание углерода и перевод его в органические соединения производится главным образом растениями и водорослями путем вовлечения в биосинтетические процессы СО2 из атмосферы. Накопленное этими организмами органическое вещество затем перерабатывается на разных уровнях жизни консументами и редуцентами. К первым принадлежат в основном животные, ко вторым – грибы и бактерии. Последовательность этих событий выражается в трофических цепях, или цепях питания. Конечное, деструктивное, звено этой цепи – минерализация органических веществ с возвратом СО2 в атмосферу – осуществляется в почвенном покрове Земли за счет деятельности гетеротрофных микроорганизмов. Около 90% образующейся из органических веществ углекислоты «микробного происхождения» и только 10% приходится на долю дыхания высших организмов и деятельности человека. При этом в аэробных условиях грибы дают 2/3, а бактерии 1/3 СО2. Часть органического вещества закрепляется в почве в форме гумуса.
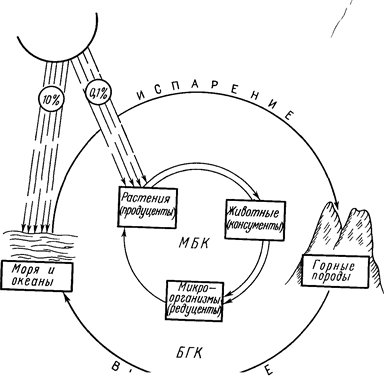
Рис. 1. Большой геологический и малый биологический круговороты
Помимо глобального процесса круговорота углерода, состоящего из синтеза и минерализации органических веществ, почва участвует в обмене многими газами с атмосферой. В ходе фотосинтетической деятельности происходит не только связывание растениями СО2, но и обогащение атмосферы кислородом. Весь кислород в газовой оболочке Земли — продукт фотосинтеза. При минерализации органических веществ в атмосферу возвращаются помимо СО2 еще СН4, H2, CO, H2S, NO2,N2 (рис.2). В почве образуются и физиологически активные летучие соединения, такие как этилен, этиламин, нитрозамин, метилртуть. Газы, образующиеся в анаэробной зоне, частично перехватываются аэробами, которые составляют «бактериальный фильтр», а частично попадают в атмосферу, откуда снова вовлекаются в круговорот. Азот из атмосферы фиксируется (биологически закрепляется) почвенными микроорганизмами. Таким образом, газы являются переносчиками энергии из анаэробной зоны в аэробную.
Превращения органических веществ и обмен газообразных продуктов микробного метаболизма сопровождается взаимодействием почвенных микроорганизмов с первичными и вторичными минералами почвы. По своему значению для биосферы этот процесс сопоставим с фотосинтезом и фиксацией молекулярного азота, так как минеральные элементы, первоисточник которых находится в литосфере, необходимы для жизни всех организмов на Земле. В процессе почвообразования происходит разрушение минералов породы и извлечение элементов, которые поступают в обменные реакции биосинтеза. Без снабжения растений из почвы такими элементами, как фосфор и калий, поступающими из минералов, было бы невозможно создание первичной растительной продукции. Разрушение минералов происходит частично под влиянием корневых систем растений, но в наибольшей степени оно осуществляется в результате жизнедеятельности почвенных микроорганизмов, которые образуют органические и минеральные кислоты, щелочи, а также выделяют во внешнюю среду синтезированные ими ферменты, полисахариды, фенольные соединения. Эти вещества прямо или косвенно взаимодействуют с минералами, разрушая кристаллические решетки, образуя комплексные соединения, переводя элементы из одной формы в другую с изменением валентности и подвижности
Таким образом, роль почвенных организмов проявляется не только в деструкции органической массы растений и животных, но также в контроле газового состава атмосферы и преобразовании литосферы, граничащей с почвой.
Высокая активность и огромные масштабы совершаемых микроорганизмами планетарных превращений веществ обусловлены их огромной численностью, повсеместным распространением, необычайной скоростью роста и разнообразием метаболических процессов. Есть микроорганизмы, способные переходить от одного типа питания к другому, например, от фотоавтотрофии к хемогетеротрофии. Такие организмы называют миксотрофами. К ним можно отнести, например, зеленую водоросль Chlorella vulgaris, которая может расти как на свету, так и в темноте за счет фотосинтеза или гетеротрофно. Миксотрофами являются и водородные бактерии. Автотрофные микроорганизмы С.Н. Виноградский назвал хемосинтезирующими, а процесс окисления неорганических соединений для получение энергии — аноргоксидацией. Деление на авто- и гетеротрофов используется в микробиологии наиболее часто. Катаболические процессы (реакции получения энергии) — это отнятие и перенос водорода (электрона). В зависимости от конечного акцептора водорода, образующегося при дегидрировании, различают следующие энергетические процессы у микроорганизмов: 1) дыхание (перенос на кислород); 2) брожение (перенос на органические акцепторы); 3) анаэробное дыхание (перенос на неорганические акцепторы — сульфаты, нитраты, хроматы). У бактерий есть также фотосинтетическое фосфорилирование - образование АТФ за счет световой энергии.
Анаболические процессы, или конструктивный (строительный) метаболизм, связаны главным образом с источниками углерода и азота, которые могут быть в органической и неорганической форме, тогда как все другие элементы микроорганизмы получают в виде минеральных солей. Соотношение С : N в биомассе бактерий составляет 5:1, а в среде оптимальное соотношение должно быть 25 : 1, так как 1/5 часть углерода включается в вещества клеток, а 4/5 расходуется на энергетические нужды. Источниками углерода служат СО2 и (или) органические соединения. В качестве источников азота выступают белки, пептиды, аминокислоты, нитраты, аммонийные соединения и молекулярный азот. Кислород и водород поступают из воды. Кроме источников основных элементов-органогенов, зольных элементов и микроэлементов многие микроорганизмы нуждаются в специфических веществах, которые носят название факторов роста. Такими факторами роста у микроорганизмов могут быть витамины, аминокислоты, пуриновые основания. Независимые от факторов роста микроорганизмы принято называть прототрофами, а их мутанты, нуждающиеся в каком-либо ростовом факторе, — ауксотрофами. Ауксотрофные мутанты широко используются в генетических исследованиях, где потребность в факторе роста служит маркером для изучения скрещивания и расщеплений в потомстве.
Специфические потребности микроорганизмов в источниках питания, факторах роста или микроэлементах делают их удобными объектами для использования в качестве биологических индикаторов, например, при определении потребности почвы в удобрениях, при количественном анализе содержания витаминов или микроэлементов в почве. Впервые на свойства микроорганизмов как химических реактивов обратил внимание В. Л. Омелянский. В 1906 г. он написал статью «О применении бактериологического метода при химическом исследовании», в которой отметил две особенности микроорганизмов как биологических индикаторов, их высокую специфичность и исключительную чувствительность. Все процессы катаболизма и анаболизма протекают не самопроизвольно, а катализируются особыми ферментами.
Ферменты есть конститутивные, т. е. всегда присущие организму независимо от условий его роста, и индуцибельные, которые синтезируются клеткой только в присутствии соответствующего субстрата. B этом одна из особенностей ферментных систем микроорганизмов. Другая особенность — образование экзоцеллюлярных (внеклеточных ) ферментов, которые расщепляют сложные молекулы субстрата (целлюлозу, белки, лигнин, хитин, липиды, пектин) во внешней среде. При этом клетка может синтезировать много фермента и поддерживать высокую активность процесса превращения экзогенного субстрата. Результатом является переработка больших количеств разнообразных веществ микроорганизмами при малой их биомассе. Эта особенность микроорганизмов широко используется человеком в таких процессах, как, например, пектиновое брожение, происходящее при мочке прядильных растений.
ПРЕВРАЩЕНИЯ СОЕДИНЕНИЙ УГЛЕРОДА
И КРУГОВОРОТ КИСЛОРОДА
В круговороте углерода и кислорода находит наиболее явное выражение взаимная зависимость и связь всех живых существ на Земле. Благодаря участию в этом цикле микроорганизмов как деструктивного звена происходит замыкание круговорота и совершается циклическое превращение веществ и энергии в биосфере. В этом — главнейшая функция микроорганизмов, которая проявляется в основном в почве.
В цикле углерода можно выделить два важнейших звена, имеющих планетарные масштабы и связанных с выделением и поглощением кислорода: 1) фиксация СО2 в процессе фотосинтеза; 2) минерализация органических веществ с выделением СО2. Первый процесс совершается в основном растениями и обеспечивает выделение кислорода в атмосферу. Второй производится микроорганизмами и идет, как правило, с поглощением кислорода (рис. 4).
ПРЕВРАЩЕНИЯ ОДНОУГЛЕРОДНЫХ СОЕДИНЕНИЙ
Из одноуглеродных соединений наибольшее значение на нашей планете имеет СО2, так как из углекислоты создается вся первичная органическая продукция. В атмосфере содержится около 0,03% СО2; в почвенном воздухе — на порядок больше. Биологическое связывание углекислоты происходит в процессах фотосинтеза, хемосинтеза гетеротрофной фиксации. Фотосинтез идет в основном в наземном слое биосферы, два последних процесса — преимущественно в почве.
Фотосинтез. Фиксация СО2 в фотосинтезе происходит за счет световой энергии СО2+Н2О+hv - (СН2О)n+О2. К фотосинтетикам принадлежат высшие растения, водоросли, пурпурные и зеленые бактерии, цианобактерии Указанное выше уравнение справедливо для растений, водорослей и цианобактерий. Пурпурные и зеленые бактерии в процессе фотосинтеза кислород не выделяют. Донором электрона у них служит не молекула воды, а восстановленные соединения серы, молекулярный водород или органическое вещество (в анаэробных условиях).
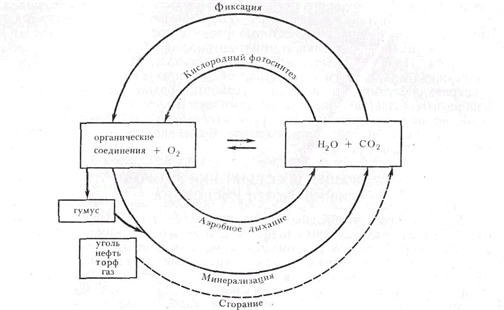
Рис 4 Круговорот углерода и кислорода
Пурпурные и зеленые серобактерии — обитатели пресных и соленых водоемов, содержащих сероводород. Это в основном водоемы застойного типа, заболоченные, где идет разложение органических (белковых) веществ в анаэробных условиях. Фотосинтезирующие серные бактерии редко можно обнаружить и в почве. Пурпурные несерные бактерии, например Rhodomicrobium, чаще выделяются из почв. Морфологически это почкующиеся бактерии, сходные с почвенными бесцветными Hyphomicrobium, но имеющие пигменты. Несерные фотосинтезирующие бактерии — анаэробы, использующие органическое вещество в качестве донора электрона. Они способны к фотоассимиляции органических субстратов (фотогетеротрофы). В эволюционном плане это, по-видимому, наиболее древние фотосинтетики. Следующий этап эволюции фотосинтеза — появление серных бактерий, которые оказались способными к гетеротрофной, а затем и автотрофной фиксации СО2 с восстановленными донорами электрона в виде H2S. Позже донором электрона в фотосинтетическом восстановлении СО2 стала служить молекула воды, а фотосинтез оказался сопряженным с выделением свободного кислорода (цианобактерии, водоросли, высшие растения). Этот тип фотосинтеза, независимый от присутствия органических источников углерода и от доноров водорода, имеющихся локально в среде, получил преимущество, что привело к доминирующему положению растений на суше, а водорослей — в океане, среди фотосинтезирующих организмов. Фиксация СО2 у всех автотрофных организмов происходит за счет универсального механизма, который известен как цикл Кальвина.
Хемосинтез. Фиксация СО2 в хемосинтезе происходит за счет энергии окисления внешнего неорганического донора электрона. Этот процесс был описан впервые С. Н. Виноградским в конце прошлого века и назван аноргоксидацией. Хемоавтотрофы известны только среди бактерий. К хемосинтезирующим организмам принадлежат нитрификаторы, карбоксидобактерии, серобактерии, тионовые, железобактерии, водородные бактерии. Они называются так по субстратам окисления, которыми могут быть NН2, NO2, CO, H2S, S, Fe2+ , Н2. Некоторые виды — облигатные хемолитоавтотрофы, другие — факультативные. К последним относятся карбоксидобактерии и водородные бактерии. В отсутствие органических веществ они используют энергию окисленного водорода или СО для фиксации СО2, а при наличии органических соединений переходят на гетеротрофный тип питания, т.е. ведут себя как миксотрофы.
Гетеротрофная фиксация СО2 . Помимо фото- и хемосинтеза, где углекислота служит единственным источником углерода, СО2 может ассимилироваться гетеротрофно, путем присоединения к готовым углеродным цепям. К гетеротрофному связыванию углекислот способны и такие фотоавтотрофы, как высшие растения. У микроорганизмов эта способность впервые была обнаружена А. Ф. Лебедевым (1921) при работе с грибами. Акцепторами СО2 могут быть различные органические кислоты, чаще всего пировиноградная:
СНзСОСООН + СО2 «СООНСН2СОСООН - щавелево-уксусная кислота.
Более 10% биомассы микроорганизмов может быть из СО2, как этобыло показано в опытах с дрожжами при использовании меченной по углероду 14СО2.
Образование метана биологическим путем из СО2 и других одноуглеродных источников — метановое брожение — происходит в болотах, в иловых отложениях озер, в метантэнках, в рубце жвачных животных. В почве этот процесс протекает там, где складываются анаэробные условия, и образуется водород в первичных процессах анаэробного превращения органических веществ. Метанобразующие бактерии — вторичные анаэробы, так как они перерабатывают продукты поступающие от других анаэробов. Метан образуется из ацетата или за счет восстановления СО2, СО, муравьиной кислоты или метанола водородом по следующей схеме СО2+4Н2 ®СН4+2Н20. Примерно 1/3 природного метана образуется из СО2. Один из наиболее известных возбудителей метанового брожения — Methanosarcina barkeri. Этот микроорганизм образует конгломераты клеток с газовыми вакуолями, что придает ему свойство всплывать на поверхность при заполнении вакуолей. После «стравливания» метана клетки увеличивают плотность и опускаются на дно в анаэробную зону, где снова осуществляют процесс образования метана. Группа метанобразующих бактерий гетерогенна. Среди них есть кокковидные, палочковидные, сарциноподобные организмы и спириллы. Все метаногенные бактерии относятся к подцарству архебактерий среди прокариот.
Окисление метана — биологический процесс, который активно протекает в районах метаногенеза — в местах газовых и нефтяных месторождений, в переувлажненных почвах, в поверхностной пленке болот. Метанокисляющие бактерии перехватывают метан, который образуется в анаэробной зоне, и окисляют его до СО2 через метанол, формальдегид и муравьиную кислоту: СН4 ® СН3ОН ® СНОН ® НСООН ® СО2. Этот путь диссимиляции (катаболизма) осуществляется с участием ферментов цепи переноса электрона. Ассимиляция углерода метана происходит на уровне формальдегида несколькими путями, отличными от автотрофной фиксации СО2 в цикле Кальвина. Микроорганизмы, окисляющие метан, получили название метилотрофов. Среди них есть облигатные формы, использующие в качестве углеродного источника питания метан или его производные — метанол и метиламины, и факультативные метилотрофы, ассимилирующие как одноуглеродные соединения (метанол, формальдегид, метиламин, но не метан), так и другие органические вещества. Облигатные метилотрофы — группа метанокисляющих бактерий сем. Methylomonadaceae, которые относят к 4 родам: Methylomonas, Methylococcus, Methylosinus и Methylocystis. Хотя формально их следует считать гетеротрофами (метан — органическое вещество), но по многим морфологическим и физиологическим признакам они близки к таким литоавтотрофам, как нитрифицирующие бактерии сем. Nitrobacteriaceae. Процесс окисления метана сопровождается у них окислением аммиака до нитритов, они имеют хорошо развитую систему внутриклеточных цитоплазматических мембран, пути метаболизма метанокисляющих бактерий сходны с таковыми у автотрофных нитратных бактерий.
Среди факультативных метилотрофов известны не только прокариоты, но и представители одноклеточных грибов. Метилотрофные дрожжи относятся к двум близким родам Pichia и Hansenula (сумчатые из Ascomycetes) или же к несовершенным грибам из рода Candida. Наиболее хорошо изученный вид С. boidinii. Он активно растет на метаноле и используется в промышленности для получения микробного белка.
Микроорганизмы-метилотрофы, особенно метанокисляющие бактерии, вовлекая в биотический круговорот одноуглеродные соединения группы метана, вносят значительный вклад в глобальный цикл углерода, замыкая трофические цепи в специфических нишах биосферы. Они представляют интерес и в практическом отношении как продуценты белка из дешевого сырья, а также как средство борьбы с метаном в угольных шахтах.
Окисление окиси углерода микроорганизмами — процесс, благодаря которому поддерживается природное равновесие этого газе в биосфере; СО образуется техногенным путем в результате сгорание разных видов топлива. Существуют и природные источники угарного газа — извержения вулканов и биохимические реакции фотосинтеза, приводящие к распаду порфиринового кольца хлорофилла. Удаление окиси углерода из атмосферы происходит в результате ее поглощения почвой и окисления микроорганизмами. Ежегодно почва поглощает до 4,1´1014 г СО, что лишь немногим меньше, чем образуется при сгорании топлива. В окислении СО участвует физиологическая группа карбоксидобактерий. Среди последних много новых, недавно открытых бактерий: Seliberia carboxydohydrogenia, Pseudomonas gazotropha Comamonas compransoris, Achromobacier carboxydus. Карбоксидобактерии — автотрофы. Образующаяся в результате окисления СО углекислота фиксируется в процессах анаболизма автотрофно через цикл Кальвина, а не метилотрофно. Но автотрофия карбоксидобактерий - облигатная. Они способны и к органотрофному росту, в том числе на метаноле. Однако высокие концентрации органических веществ подавляют их развитие, на основании чего их относят к олиготрофной группировке почв. Г. А. Заварзин (1978) относит их к микрофлоре рассеяния.
ПРЕВРАЩЕНИЯ БЕЗАЗОТИСТЫХ ОРГАНИЧЕСКИХ
СОЕДИНЕНИЙ
Основной источник пополнения запасов углекислоты в атмосфере — распад растительных тканей с высоким отношением С : N. Если бы не постоянно идущие процессы минерализации, производимые почвенными микроорганизмами, вся углекислота за несколько десятков лет могла бы быть исчерпана на фотосинтез. Органические вещества, входящие в состав растительных тканей, можно по степени их сложности и доступности для микробного разложения, разделить на следующие группы:
1) растворимые в воде соединения — сахара, органические кислоты, спирты;
2) слаборастворимые — гемицеллюлозы;
3) нерастворимые — крахмал, пектин, клетчатка, воска, жиры, смолы, углеводороды;
4) особо прочное соединение — лигнин.
Рассмотрим разложение этих веществ, последовательно по группам, в разных природных условиях — аэробных и анаэробных.
Разложение водорастворимых органических соединений происходит в первую очередь после отмирания растений. Потеря углеводов приводит к сужению отношения С : N в разлагающемся материале и торможению процесса распада. В аэробных условиях углеводы разлагаются бактериями в актах дыхания, с образованием СО2 как конечного продукта деструкции. Грибы обычно окисляю углеводы не до конца, а процесс идет с образованием недоокисленных продуктов, в первую очередь органических кислот: янтарной, лимонной, уксусной и др. Органические кислоты в большом количестве образуются при анаэробном распаде углеводов — брожении.
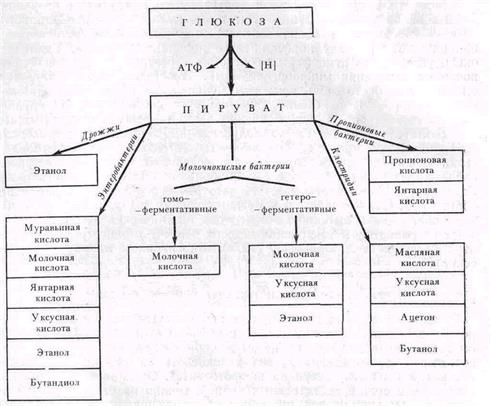
Рис 5 Типы брожений
В зависимости от возбудителей и конечных продуктов различают несколько типов брожений (рис. 5). При сбраживании глюкозы дрожжами образуется этиловый спирт, при молочнокислом брожении главным или единственным продуктом является молочная кислота. Клостридии сбраживают глюкозу до масляной кислоты, пропионовые лактобациллы — до пропионовой и янтарной. Спиртовое брожение
!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!
Изучение процессов разложения клетчатки в природе было начато еще в прошлом веке под влиянием идей Пастера об анаэробных брожениях. Поэтому в первую очередь изучали анаэробное разложение целлюлозы. Биологическую природу этого процесса впервые установил Л. Попов в 1875 г. Позже В. Л. Омелянский подробно изучал анаэробный распад клетчатки и выделил возбудителей этого процесса, один из которых был назван в его честь Bacillus omelianskii. Это тонкие, до 12 мкм в длину, слегка изогнутые палочки с округлыми или овальными терминальными спорами («барабанные палочки»). При анаэробном разложении целлюлозы образуется много органических кислот (уксусная, янтарная, молочная, масляная, муравьиная), этиловый спирт, СО2 и водород. Поэтому анаэробное разложение целлюлозы сопровождается активным развитием сопутствующих микроорганизмов, часто азотфиксаторов. Сбраживание клетчатки происходит в природе и в условиях повышенных температур, например при разогревании торфа, соломы, компостов. В этом случае основной возбудитель процесса — термофильная бактерия Clostridium ihermocellum. Выделенный из горячих источников анаэроб Thermoanaerobacter efhanolicus при сбраживании клетчатки образует большое количество этанола. Он использует кроме целлюлозы и продукты ее распада — целлобиозу и глюкозу, в отличие от Clostridium ihermocellum, рост которого подавляется продуктами распада целлюлозы. Их совместное культивирование дает возможность из целлюлозного сырья получать этиловый спирт путем экономичной ферментации при высоких температурах. Другая термофильная бактерия — Thermoanaefobium brockii не использует целлюлозу, но хорошо растет на целлобиозе, также продуцируя этанол. Этот микроорганизм выделяется из горячих источников вулканического происхождения и имеет температурный диапазон роста между 35 и 85° с оптимумом 65—70°.
В отличие от процесса анаэробного разложения целлюлозы, который осуществляется только бактериями, в аэробных условиях клетчатку разлагают многие микроорганизмы самых разных систематических групп: истинные бактерии, миксобактерии, актиномицеты и грибы.
В кислых лесных почвах, где клетчатка в значительной степени представлена мертвой древесиной, она более доступна для грибного разложения. Поэтому в лесах главная роль в превращении целлюлозы принадлежит грибам, в особенности почвенным микромицетам. Среди них наиболее активны такие типичные подстилочные сапрофиты, как Trichoderma viride, Chaetomium globosum, Myrothecium verrucaria, а также некоторые виды родов Penicillium и Aspergillus. Характерная особенность грибного разложения целлюлозы — выделение целлюлазных ферментов в среду. Поэтому грибы представляют промышленный интерес для получения препарата целлюлаз после освобождения среды от мицелия. У бактерий целлюлазы — контактные ферменты, связанные, по-видимому, клеточной поверхностью. Они работают только in situ, и выделение ферментов из этих микроорганизмов представляет определенные трудности.
В почвах под травянистой растительностью — в степных и луговых ландшафтах — разложение целлюлозы осуществляют главным образом миксобактерии и цитофаги, а также истинные бактерии и актиномицеты.
Жиры входят в состав всех растительных и животных тканей. Некоторые органы растений, например семена, бывают очень богаты жирами. В почве, обогащенной гумусом и растительными остатками, липидная фракция может составлять 3% и более. Жиры разлагаются в почве микроорганизмами, обладающими липазной активностью. При разложении жиров образуются глицерин и жирные кислоты. В аэробных условиях глицерин быстро используется в метаболизме многих бактерий и грибов, а жирные кислоты более стойки и они накапливаются в почве. В анаэробных условиях жирные кислоты восстанавливаются до углеводородов. Липазы имеются у аэробных почвенных бактерий, актиномицетов и грибов, а также у анаэробов из рода Clostridium.
Углеводороды в почве представлены газообразными (метан, этан, этилен и др.), алифатическими и циклическими соединениями. Об окислении метана говорилось выше. Алифатические углеводороды с длинной цепью окисляют многие бактерии и дрожжи. Из бактерий наиболее активны представители коринеформной группы, псевдомонады и нокардии, из дрожжей — виды родов Candida, Debaryomyces, Schwanniomyces. Большое значение имеет длина углеродной цепи: легче используются углеводороды, начиная с гексадекана. Утилизация углеводородов происходит в окислительных условиях, так как первый этап воздействия на углеводородную цепь — окисление концевого углерода ферментами оксигеназами. Дальнейшее окисление протекает по пути, который известен как окисление жирных кислот. При низком парциальном давлении кислорода происходит накопление промежуточных продуктов окисления углеводородов — жирных кислот, начиная с Сз (пропионовой кислоты).
Разложение микроорганизмами ароматических углеводородов в почве имеет очень большое значение в круговороте углерода, так как эти соединения входят в состав молекул лигнина. Способность разлагать ароматические углеводороды присуща многим грибам, некоторым аэробным бактериям и дрожжам. Из бактерий наиболее активны представители родов Pseudomonas и Arthrobacter. В анаэробных условиях ароматические соединения могут ассимилироваться (только на свету!) некоторыми фототрофными бактериями. Разложение циклических углеводородов начинается с гидроксилирования — включения в них оксигрупп и перевода в орто - или парадиоксифенилпроизводные. Далее следует разрыв кольца под действием оксигеназ, катализирующих окисление субстрата кислородом. Боковые цепи и заместители отщепляются, как правило, до разрыва кольца. Разрыв ароматического кольца происходит разными путями у разных микроорганизмов. Из почв были выделены бактерии, разлагающие полициклические соединения углеводородной природы — нафталин, антрацен и фенантрен. Промежуточным продуктом их распада является салициловая кислота.
Итак, большинство природных углеводородов частично или полностью окисляются микроорганизмами. Этот процесс важен в аспекте самоочищения почвы от загрязнений нефтью и продуктами ее переработки.
Лигнин (от лат. lignum — дерево, древесина) синтезируется только высшими растениями и в количественном отношении уступает только целлюлозе и гемицеллюлозам. В древесине лиственных пород деревьев его содержится 20—30%, хвойных — до 50%. Лигнин относится к инкрустирующим веществам клеточной стенки. Он вызывает одревеснение клеток и придает им большую прочность. Если целлюлозные микрофибриллы сравнить с арматурой железобетонных конструкций, то лигнину следует отвести роль бетона. Химическое строение лигнина окончательно не установлено. Известно, что в разных растениях он неоднороден. Сложность структуры определяется большим числом полимеризованных мономерных блоков, которые представляют собой производные фенилпропана. Основной мономер лигнина — конифериловый спирт; он составляет главный скелет лигнина хвойных. В лигнине лиственных пород есть еще синаповый спирт, а в лигнине из соломы злаков — кумаровый.
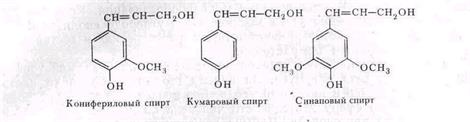
Лигнин не растворяется ни в воде, ни в органических растворителях» дает цветные реакции, характерные для фенолов. В растительном организме — это конечный продукт обмена, выполняющий только механические функции. Разлагают его грибы и немногие бактерии. Процесс протекает очень медленно, поэтому промежуточные продукты разложения лигнина накапливаются в почве. Основные разрушители лигнина — базидиальные грибы. Некоторые из них разрушают лигнин даже в живых растениях, например осенний опенок Armillariella mellea. Древоразрушающие базидиомицеты, вызывающие белую гниль дерева, наиболее активные разрушители лигнина вместе с некоторыми подстилочными сапрофитами. Можно проследить сукцессию видов при разложении лигнина. Процесс начинается еще в растущем дереве, на котором поселяются базидиомицеты — паразиты родов Fomes, Polyporus, Armillariella и др. На мертвой древесине появляется другая группировка грибов: это деревоокрашивающие грибы Ceraiostomella, Cladosporium. Далее частично разрушенную древесину заселяют подстилочные сапрофиты — базидиомицеты с мелкими шляпочными плодовыми телами родов Collybia, Marasmius, Phallus, Lepiota, Mycena и, наконец, в поздней стадии происходит колонизация остатков почвенными грибами из группы несовершенных: Fusarium, Trichoderma, Aspergillus, Penicillium, Rhizoclonia. В более поздних стадиях разложения лигнина участвуют и бактерии, причем только грамотрицательные. Древесина и опад лиственных деревьев разрушаются быстрее, чем хвойных. Разложение лигнина идет обычно параллельно с распадом целлюлозы. Оба эти процесса могут вызывать одни и те же организмы. Например, у Polyporus abiciinus найдено до двух десятков внеклеточных ферментов, среди которых есть и целлюлазы, и полифенолоксидазы, разлагающие лигнин, и, кроме того, ферментативный комплекс, воздействующий на пектины, крахмал, гемицеллюлозы. Несомненно, что в природе разложение лигнина совершается сложным комплексом микроорганизмов и высших грибов, и этот процесс идет значительно интенсивнее, чем при лабораторных анализах с чистыми культурами
ЗАХОРОНЕННЫЙ УГЛЕРОД И ЕГО МОБИЛИЗАЦИЯ
Часть углерода на нашей планете закре
Дата добавления: 2020-10-01; просмотров: 1063;
Поиск по сайту
Узнать еще
- III. Контроль эффективности
- III.4. Геоэкологическая роль технического прогресса
- IX.2. Биотическое управление экосферой и роль деятельности человека
- S Власть и контроль
- V. Контроль за корректурой СККРиПДП.
- VII. ОБСЛЕДОВАНИЕ ПОМЕТОВ И КОНТРОЛЬ КАЧЕСТВА ИХ ВЫРАЩИВАНИЯ
- VIII. 2. Большой круговорот вещества и роль в нем человека
- Інфраструктура: сутність і роль у механізмі функціонування національного ринку

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
