Эмбриональный период
Дробление
Дробление — это ряд последовательных митотических делений зиготы, в результате которых огромный объем цитоплазмы яйца разделяется на многочисленные, содержащие ядра клетки меньшего размера. В результате дробления образуются клетки, которые называют бластомерами. Дробление от обычного деления отличает то, что вновь образовавшиеся бластомеры не увеличиваются в размерах. Это становится возможным вследствие выпадения пресинтетического периода интерфазы. При этом синтетический период интерфазы начинается в телофазе предшествующего митоза. Таким образом, количество бластомеров постепенно увеличивается, а их общий объем практически не изменяется. Цитоплазма клеток при дроблении делится путем возникновения впячиваний оболочки клетки (борозды дробления).
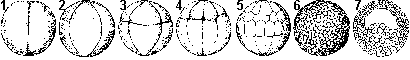
Дробление яйцеклетки амфибий (лягушка): 1 — двуклеточная стадия; 2 — четырехклеточная стадия; 3 — восьмиклеточная стадия;
4 — переход от восьми- к шестнадцатиклеточной стадии (клетки анимального полюса уже поделились, а клетки вегетативного
только начинают дробиться); 5 — более поздняя стадия дробления; 6 — бластула; 7 — бластула в разрезе.
Биологическое значение процесса дробления: благодаря повторяющимся циклам репродукции, происходит размножение генотипа зиготы; происходит накопление клеточной массы для дальнейших преобразований, зародыш из одноклеточного превращается в многоклеточный.
Деление бластомеров бывает синхронным и несинхронным. У большинства видов оно несинхронно с самого начала развития, у других становится таковым уже после первых делений.
Характер дробления определяется, прежде всего, строением яйцеклетки, главным образом, количеством желтка и особенностями его распределения в цитоплазме. В этой связи по способу дробления выделяют два основных типа яиц: полностью дробящиеся и дробящиеся частично. Полным дробление называется тогда, когда цитоплазма яйцеклетки полностью разделяется на бластомеры. Оно может быть равномерным — все образовавшиеся бластомеры имеют одинаковые размеры и форму (характерно для алецитальных и изолецитальных яйцеклеток) и неравномерным — образуются неравные по размерам бластомеры (свойственно телолецитальным яйцеклеткам с умеренным содержанием желтка). Мелкие бластомеры возникают у анимального полюса, крупные — в области вегетативного полюса зародыша.
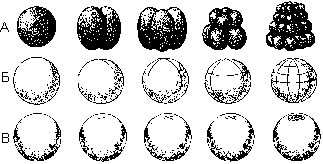
Различные виды дробления: А — полное; Б — частичное; В — дискоидальное.
Частичное дробление — тип дробления, при котором цитоплазма яйцеклетки не полностью разделяется на бластомеры. Одним из видов частичного дробления является дискоидальное, при котором дроблению подвергается только лишенный желтка участок цитоплазмы у анимального полюса, где находится ядро. Участок цитоплазмы, подвергшийся дроблению, называется зародышевым диском. Этот тип дробления характерен для резко телолецитальных яиц с большим количеством желтка (рептилии, птицы, рыбы).
Дробление у представителей разных групп животных имеет свои особенности, однако завершается оно образованием близкой по строению структуры — бластулы.
Бластула
Бластула — однослойный зародыш. Она состоит из слоя клеток — бластодермы, ограничивающей полость — бластоцель. Бластула начинает формироваться на ранних этапах дробления благодаря расхождению бластомеров. Возникающая при этом полость заполняется жидкостью. Строение бластулы во многом зависит от типа дробления.
Целобластула (типичная бластула) образуется при равномерном дроблении. Имеет вид однослойного пузырька с большим бластоцелем (ланцетник).
Амфибластула образуется при дроблении телолецитальных яиц; бластодерма построена из бластомеров разного размера: микромеров на анимальном и макромеров на вегетативном полюсах. Бластоцель при этом смещается в сторону анимального полюса (земноводные).
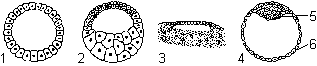
Типы бластул: 1 — целобластула; 2 — амфибластула; 3 — дискобластула; 4 — бластоциста; 5 — эмбриобласт; 6 — трофобласт.
Дискобластула образуется при дискоидальном дроблении. Полость бластулы имеет вид узкой щели, находящейся под зародышевым диском (птицы).
Бластоциста представляет собой однослойный пузырек, заполненный жидкостью, в котором различают эмбриобласт (из него развивается зародыш) и трофобласт, обеспечивающий питание зародыша (млекопитающие).
Гаструла
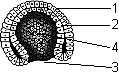
Гаструла:
1 — эктодерма; 2 — энтодерма; 3 — бластопор; 4 — гастроцель.
После того как сформировалась бластула, начинается следующий этап эмбриогенеза — гаструляция (образование зародышевых листков). В результате гаструляции образуется двухслойный, а затем трехслойный зародыш (у большинства животных) — гаструла. Первоначально образуются наружный (эктодерма) и внутренний (энтодерма) слои. Позже между экто- и энтодермой закладывается третий зародышевый листок — мезодерма.
Зародышевые листки — отдельные пласты клеток, занимающие определенное положение в зародыше и дающие начало соответствующим органам и системам органов. Зародышевые листки возникают не только в результате перемещения клеточных масс, но и в результате дифференциации сходных между собой, сравнительно однородных клеток бластулы. В процессе гаструляции зародышевые листки занимают положение, соответствующее плану строения взрослого организма. Дифференциация — процесс появления и нарастания морфологических и функциональных различий между отдельными клетками и частями зародыша. В зависимости от типа бластулы и от особенностей перемещения клеток, различают следующие основные способы гаструляции: инвагинация, иммиграция, деламинация, эпиболия.
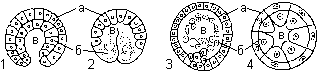
Типы гаструл: 1 — инвагинационная; 2 — эпиболическая; 3 — иммиграционная; 4 — деламинационная;
а — эктодерма; б — энтодерма; в — гастроцель.
При инвагинации один из участков бластодермы начинает впячиваться внутрь бластоцеля (у ланцетника). При этом бластоцель практически полностью вытесняется. Образуется двухслойный мешок, наружная стенка которого является первичной эктодермой, а внутренняя — первичной энтодермой, выстилающей полость первичной кишки, или гастроцель. Отверстие, при помощи которого полость сообщается с окружающей средой, называется бластопором, или первичным ртом. У представителей разных групп животных судьба бластопора различна. У первичноротых животных он превращается в ротовое отверстие. У вторичноротых бластопор зарастает, и на его месте нередко возникает анальное отверстие, а ротовое отверстие прорывается на противоположном полюсе (переднем конце тела).
Иммиграция — «выселение» части клеток бластодермы в полость бластоцеля (высшие позвоночные). Из этих клеток образуется энтодерма.
Деламинация встречается у животных, имеющих бластулу без бластоцеля (птицы). При таком способе гаструляции клеточные перемещения минимальны или совсем отсутствуют, так как происходит расслоение — наружные клетки бластулы преобразуются в эктодерму, а внутренние формируют энтодерму.
Эпиболия происходит, когда более мелкие бластомеры анимального полюса дробятся быстрее и обрастают более крупные бластомеры вегетативного полюса, образуя эктодерму (земноводные). Клетки вегетативного полюса дают начало внутреннему зародышевому листку — энтодерме.
Описанные способы гаструляции редко встречаются в чистом виде и обычно наблюдаются их сочетания (инвагинация с эпиболией у амфибий или деламинация с иммиграцией у иглокожих).
Чаще всего клеточный материал мезодермы входит в состав энтодермы. Он впячивается в бластоцель в виде карманообразных выростов, которые затем отшнуровываются. При образовании мезодермы происходит образование вторичной полости тела, или целома.
Процесс формирования органов в эмбриональном развитии называют органогенезом. В органогенезе можно выделить две фазы: нейруляция — образование комплекса осевых органов (нервная трубка, хорда, кишечная трубка и мезодерма сомитов), в который вовлекается почти весь зародыш, и построение остальных органов, приобретение различными участками тела типичной для них формы и черт внутренней организации, установление определенных пропорций (пространственно ограниченные процессы).
По теории зародышевых листков Карла Бэра, возникновение органов обусловлено преобразованием того или иного зародышевого листка — экто-, мезо- или энтодермы. Некоторые органы могут иметь смешанное происхождение, то есть они образованы при участии сразу нескольких зародышевых листков. Например, мускулатура пищеварительного тракта является производным мезодермы, а его внутренняя выстилка — производное энтодермы. Однако, несколько упрощая, происхождение основных органов и их систем все-таки можно связать с определенными зародышевыми листками. Зародыш на стадии нейруляции называется нейрулой. Материал, используемый на построение нервной системы у позвоночных животных, — нейроэктодерма, входит в состав дорсальной части эктодермы. Он располагается над зачатком хорды.
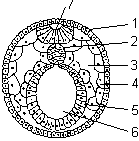
Нейрула:
1 — эктодерма; 2 — хорда; 3 — вторичная полость тела; 4 — мезодерма; 5 — энтодерма; 6 — кишечная полость; 7 — нервная трубка.
Сначала в области нейроэктодермы происходит уплощение клеточного пласта, что приводит к образованию нервной пластинки. Затем края нервной пластинки утолщаются и приподнимаются, образуя нервные валики. В центре пластинки за счет перемещения клеток по средней линии возникает нервный желобок, разделяющий зародыш на будущие правую и левую половины. Нервная пластинка начинает складываться по средней линии. Края ее соприкасаются, а затем смыкаются. В результате этих процессов возникает нервная трубка с полостью — невроцелем.
Смыкание валиков происходит сначала в средней, а затем в задней части нервного желобка. В последнюю очередь это происходит в головной части, которая по ширине превосходит другие. Передний расширенный отдел в дальнейшем образует головной мозг, остальная часть нервной трубки — спинной. В результате нервная пластинка превращается в нервную трубку, лежащую под эктодермой.
В ходе нейруляции часть клеток нервной пластинки не входит в состав нервной трубки. Они образуют ганглиозную пластинку, или нервный гребень, — скопление клеток вдоль нервной трубки. Позднее эти клетки мигрируют по всему зародышу, образуя клетки нервных узлов, мозгового вещества надпочечников, пигментные клетки и т.п.
Из материала эктодермы, помимо нервной трубки, развиваются эпидермис и его производные (перо, волосы, ногти, когти, кожные железы и т.д.), компоненты органов зрения, слуха, обоняния, эпителий ротовой полости, эмаль зубов.
Мезодермальные и энтодермальные органы формируются не после образования нервной трубки, а одновременно с ней. Вдоль боковых стенок первичной кишки путем выпячивания энтодермы образуются карманы, или складки. Участок энтодермы, расположенный между этими складками, утолщается, прогибается, сворачивается и отшнуровывается от основной массы энтодермы. Так появляется хорда. Возникшие карманообразные выпячивания энтодермы отшнуровываются от первичной кишки и превращаются в ряд сегментарно-расположенных замкнутых мешков, называемых также целомическими мешками. Их стенки образованы мезодермой, а полость внутри представляет собой вторичную полость тела (или целом).
Из мезодермы развиваются все виды соединительной ткани, дерма, скелет, поперечно-полосатая и гладкая мускулатура, кровеносная и лимфатическая системы, половая система.
Из энтодермы развиваются эпителий кишечника и желудка, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, секретирующие отделы передней и средней доли гипофиза, щитовидной и паращитовидной желез.
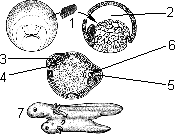
Эмбриональная индукция:
1 — зачаток хордомезодермы; 2 — полость бластулы; 3 — индуцированная нервная трубка; 4 — индуцированная хорда; 5 — первичная нервная трубка; 6 — первичная хорда; 7 — формирование вторичного зародыша, соединенного с зародышем-хозяином.
Дата добавления: 2016-05-30; просмотров: 4051;
Поиск по сайту
Узнать еще
- I. Инкубационный период
- II. ОСНОВНОЙ ПЕРИОД
- II. ПОДГОТОВИТЕЛЬНЫЙ ПЕРИОД ТРЕНИРОВКИ
- III. ОСНОВНОЙ ПЕРИОД ТРЕНИРОВКИ
- III. Периоды колонизации стран Востока
- Автоматическое управление периодическими процессами хим. очистки воды.
- АДПТАЦИЯ, ЕЕ ВИДЫ И ПЕРИОДЫ
- Амстердам. Дом-музей Рембрандта ( жил в период 1639-1658)

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
