ЖЕНСКИЕ ПОЛОВЫЕ ГОРМОНЫ
В яичниках синтезируются женские половые гормоны — эстрогены и прогестины, среди которых наиболее активны 17β-эстрадиол и прогестерон.
Синтез.
1. Превращение ХС в прегненалон катализирует в митохондриях клеток теки фолликулов холестеролдесмолаза (Р450), ее активирует через аденилатциклазную систему ЛГ. В клетках теки фолликулов синтезируются андрогены (андростендион). В клетках теки синтезируется очень небольшое количество эстрогенов.
2. В клетках гранулёзы (яичников) андрогены с участием Р450-оксидазы (требует О2 и НАДФH2) ароматизируются в эстрадиол. Р450-оксидазу в клетках гранулёзы активирует ФСГ. В клетках гранулёзы может синтезироваться менее активный эстроген— эстрон.
Значительная часть андрогенов ароматизируется в эстрогены на периферии: в жёлтом теле, фетоплацентарном комплексе (во время беременности), корой надпочечников, в жировых клетках, печени, коже и других тканях.
Малоактивный эстриол образуется из эстрона в крови.
Прогестерон выделяется главным образом жёлтым телом во время менструации (в лютеиновую фазу) и фетоплацентарным комплексом во время беременности. В небольших количествах он вырабатывается у женщин и мужчин корой надпочечников.
В фолликулярной фазе менструального цикла концентрация прогестерона в плазме обычно не превышает 5 нмоль/л, а в лютеиновой фазе увеличивается до 40—50 нмоль/л.
Секреция
Регуляция секреции
1. Из гипоталамуса импульсно секретируется гонадотропин-рилизинг-гормон (ГТГ) (декапептид).
2. Гондотропный гормон импульсно стимулирует в гипофизе синтез и секрецию фолликулостимулирующего (ФСГ) и лютеинизирующего (ЛГ) гормонов (гонадотропные гормоны гипофиза, гликопротеины, около 30 кД).
3.
· ЛГ у женщин стимулирует продукцию андрогенов клетками теки, образование прогестерона клетками жёлтого тела.
· ФСГ ускоряет развитие фолликулов в яичниках, стимулирует ароматизацию андрогенов с образованием эстрогенов.
В детском возрасте незрелые яичники вырабатывают небольшое количество гормонов, поэтому концентрация эстрогенов в крови низкая.
В пубертатный период чувствительность гипоталамо-гипофизарной системы к действию эстрогенов снижается. Импульсная секреция гонадотропин-рилизинг-гормона устанавливает суточный ритм секреции ЛГ и ФСГ.
1. В начале менструального цикла ФСГ и ЛГ вызывают развитие первичных фолликулов. Созревающий фолликул секретирует эстрогены, которые угнетают секрецию ФСГ. Яичники секретируют белок ингибин, который также тормозит секрецию ФСГ.
2. Фолликулярная фаза. Созревающий фолликул синтезирует эстрадиол, который по механизму положительной обратной связи повышает секрецию ЛГ и ФСГ.
3.Лютеиновая фаза. Повышение ЛГ приводит к овуляции — освобождению яйцеклетки из лопнувшего фолликула. После овуляции клетки гранулёзы превращаются в жёлтое тело, которое, помимо эстрадиола, начинает вырабатывать всё большее прогестерона.
При наступлении беременности, жёлтое тело продолжает функционировать и секретировать прогестерон, однако на более поздних этапах беременности прогестерон в основном продуцируется плацентой.
Если оплодотворение не происходит, высокая концентрация прогестерона в плазме крови по механизму отрицательной обратной связи угнетает активность гипоталамо-гипофизарной системы, тормозится секреция ЛГ и ФСГ, жёлтое тело разрушается, и снижается продукция стероидов яичниками. Наступает менструация, которая длится примерно 5 дней, после чего начинает формироваться новый поверхностный слой эндометрия, и возникает новый цикл.
Транспорт. Примерно 95% циркулирующих в крови эстрогенов связано с транспортными белками — СГСБ (секс-гормонсвязывающий белок) и альбумином. Биологической активностью обладает только свободная форма эстрогенов. Прогестерон в крови связывается с транспортным глобулином транскортином и альбумином, и только 2% гормона находится в свободной биологически активной форме.
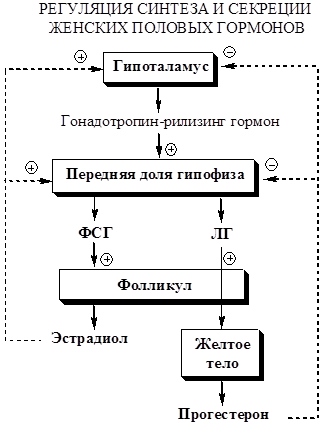
Действие гормонов
Эстрогены через ядерные рецепторы регулируют транскрипцию свыше 50 структурных генов.
Эстрогены:
1. стимулируют развитие тканей, участвующих в размножении;
2. определяют развитие женских вторичных половых признаков;
3. регулируют транскрипцию гена рецептора прогестина;
4. вместе с прогестинами в лютеиновой фазе превращают пролиферативный эндометрий (эпителий матки) в секреторный, подготавливая его к имплантации оплодотворённой яйцеклетки;
5. Совместно с простагландином F2a увеличивают чувствительность миометрия к действию окситоцина во время родов;
6. Оказывают анаболическое действие на кости и хрящи;
7. поддерживают нормальную структуру кожи и кровеносных сосудов у женщин;
8. Способствуют образованию оксида азота в сосудах гладких мышц, что вызывает их расширение и усиливает теплоотдачу.
9. Стимулируют синтез транспортных белков тиреоидных и половых гормонов.
10. Влияют на обмен липидов - увеличивают синтез ЛПВП и тормозят образования ЛПНП, что приводит к снижению содержания ХС в крови.
11. Могут индуцировать синтез факторов свёртывания крови II, VII, IX и X, уменьшать концентрацию антитромбина III.
Прогестерон:
1. влияет в основном на репродуктивную функцию организма;
2. увеличивает базальную температуру тела на 0,2-0,5 С, которое происходит сразу после овуляции и сохраняется на протяжении лютеиновой фазы менструального цикла.
3. Высокие концентрации прогестерон взаимодействует с рецепторами альдостерона почечных канальцев. В результате альдостерон теряет возможность стимулировать реабсорбцию натрия.
4. Действует на ЦНС, вызывая некоторые особенности поведения в предменструальный период.
6. Инактивация. Катаболизм половых гормонов происходит в основном в печени. При катаболизме половых гормонов, как и кортикоидов, образуются 17-окси- и 17-кетостероиды. У мужчин 2/3 кетостероидов образуется за счёт кортикостероидов и 1/3 за счёт тестостерона (всего 12—17 мг/сут). У женщин 17-кетостероиды образуются преимущественно за счёт кортикостероидов (7—12 мг/сут). В печени эстрадиол инактивируется в результате гидроксилирования ароматического кольца и образования конъюгатов с серной или глюкуроновой кислотами, которые и выводятся из организма с жёлчью или мочой. Т½ ФСГ = 150 мин, а Т½ ЛГ = 30 мин, Т½ прогестерона = 5 мин.
Дата добавления: 2016-06-29; просмотров: 2427;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
