ПРОНИКНОВЕНИЕ СТИЛЕТОВ В ТЕЛО ПИТАЮЩЕГО ОРГАНИЗМА
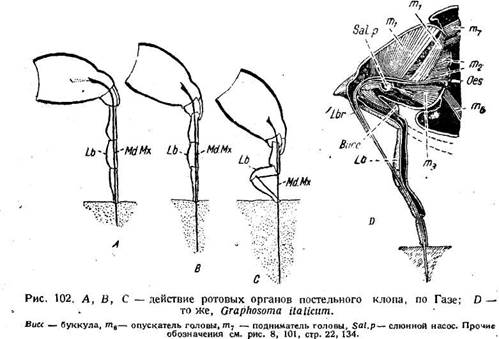
Перейдем теперь к механизму проникновения гемиптероидного хоботка в тело питающего организма. Внешняя сторона этого процесса состоит в том, что конец нижней губы фиксируется на выбранном месте покровов, после чего стилеты углубляются в тело, а нижняя губа отгибается назад коленообразно. Место сгиба может находиться в различных местах — у постельного клопа между третьим и четвертым члениками (рис. 102 С), у Graphosoma между первым и вторым (рис. 102 D). В общем процесс напоминает весьма близко работу диптероидного хоботка. У некоторых Homoptera нижняя губа не сгибается, а лишь укорачивается благодаря мягкости своего основания. Телескопическое вдвигание члеников нижней губы перед ее сгибанием происходит также и у постельного клопа (рис, 102 А, В).
Интересен вопрос о причине, заставляющей стилеты постельного клопа входить в кожу человека. На самом деле, как бы они ни былигф остры, необходим известный нажим на кожу, чтобы ее проколоть. Но при ничтожном весе насекомого (не забудем, что кусаются и личинки всех возрастов) нажим не может быть значительным. Выяснилось, однако, что при укусе клоп сперва вцепляется передними лапками в кожу и после, этого нажимает на нее хоботком, опуская голову. При ампутации передних ног, клоп становится неспособным к прокалыванию. Таким образом, вес насекомого играет лишь второстепенную роль, и прокалывание происходит за счет работы шейной и ножной мускулатуры. Не исключена, по нашему мнению, возможность, что аналогичный механизм имеет значение для действия других колющих, хоботков, например комаров. Нечто сходное имеет место также у вшей (стр. 145).
Самое проникновение стилетов в покровы, как показал Вебер, распадается на серию фаз, сгруппированных в периоды. В первой фазе концы всех четырех стилетов упираются в поверхность покровов (рис. 103,1). Во второй фазе (2) одна из мандибул прокалывает покровы, входит в их толщу на некоторую глубину и останавливается. В третьей фазе (3) друга мандибула выполняет аналогичное движение так, что концы обеих оказываются рядом в глубине покровов, но максиллы еще сохраняют прежнее положение. Наконец, в четвертой фазе (4) обе максиллы одновременно вдвигаются в толщу покровов настолько же, насколько вошли туда мандибулы. Таким образом, в четвертой фазе покровы питающего организма проколоты всеми четырьмя стилетами, и первый период прокалывания завершен. Но положение стилетов по отношению друг к другу повторяет теперь первую фазу, так как их концы находятся вместе. Поэтому четвертая фаза, завершая первый период, служит в то же время началом второго. Второй период снова начинается с продвижения одной мандибулы (5), затем ее догоняет другая (6), затем до того же уровня продвигаются максиллы (7). В конце второго периода все стилеты находятся на один «шаг» глубже, чем в конце первого. Третий период опять начинается с продвиижения одной мандибулы (8) и т.д. В результате концы стилетов погружаются все глубже и глубже.

| 
|
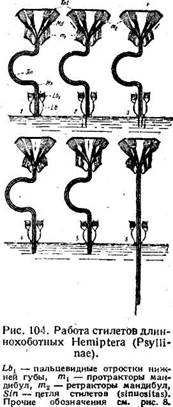
Описанная последовательность движений стилетов является, по-видимому, общей для всего отряда. Но у форм с удлиненными стилетами к ней присоединяется работа еще одного весьма замечательного механизма, раскрытого на листоблошках. На рис. 104. схематически представлены стилеты со своими мышцами и конец нижней губы. При этом обе максиллы, ввиду их синхронного действия, изображены отдельно лишь в основных своих частях, а на остальном протяжении нарисованы в виде единой полоски. Сгибание стилетов схематически представлено в виде направленной влево петли (Sin), которая отвечает как простой петле листоблошки или червеца, так и спирали Aradus. В нижней губе заключен следующий механизм. Желоб нижней губы образует два упругих пальцевидных отростка (Lb-L), которые прижимают с боков все стилеты друг к другу, и тогда движение последних или невозможно, или затруднено.
Внутри отростков имеются мышцы, при сокращении которых отростки раздвигаются и стилеты освобождаются. В первой фазе конец нижней губы и стилетов прикасаются к покровам питающего организма (рис. 104, 1). Во второй фазе сокращаются мышцы пальцевидных отростков, освобождая стилеты, которые получают возможность движения, в то же время сокращается протрактор одной из мандибул (т 1), и ее конец погружается под кожу по уже знакомой нам схеме. Чрезвычайно существенным является здесь следующий момент. Мандибулярный стилет весьма гибок, но длина его постоянна, поэтому, если при сокращении протрактора основание мандибулы смещается на некоторое расстояние, то на такое же расстояние смещается и ее дистальный конец, несмотря на то, что весь мандибулярный стилет петлеобразно изогнут. В третьей фазе (3) сокращается протрактор другой мандибулы, в четвертой (4) — максиллярные протракторы, и концы всех стилетов оказываются рядом. В результате сокращения четырех протракторов, отвечающие им четыре ретрактора (т 2) растягиваются. После этого наступает весьма важная пятая фаза (5). Она состоит, во-первых, в том, что мышцы пальцевидных отростков расслабляются и отростки автоматически прижимаются к стилетам. Во-вторых, протракторы расслабляются, а все четыре ретрактора сокращаются одновременно. При этом они тянут стилеты вверх, вытаскивая их обратно из покровов. Но вытаскивание мандибул невозможно, так как они заякорены в толще покровов своими зубцами (ср. рис. 96), а вытаскивание максилл невозможно потому, что они прижаты пальцевидными отростками к заякоренным мандибулам. Таким образом, концы всех стилетов фиксированы в достигнутом положении, а ретракторы втягивают их основания внутрь головной капсулы. При постоянной длине стилетов втягивание может осуществиться только за счет расправления петли. Это действительно и происходит, и потому в пятой фазе сгиб петли становится менее глубоким, чем в первой. Но в то же время в пятой фазе концы всех стилетов лежат на одном уровне, и потому она служит исходной для начала следующего периода. Снова разжимаются пальцевидные отростки, снова протракторы вталкивают все стилеты глубже и т. д., и петля расправляется еще в некоторой мере. Несколько таких последовательных периодов полностью расправляют даже такие образования, как петля червецов или спираль Aradus (6), и, что то же самое, вся длина стилетов выдвигается из нижней губы и проникает на относительно громадную глубину в ткани, растения или в расщелины коры, где находятся грибки.
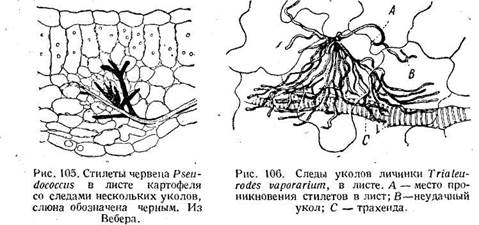
При втягивании стилетов в голову те же самые сокращения и расслабления мышц следуют в обратном порядке. Описанный механизм, однако, объясняет не все сложные движения, которые выполняются стилетами. Метод окрашивания выпущенной червецом слюны (рис. 105, 106) показывает, что стилеты, будучи введены в лист, не остаются на одном месте, но многократно колют в самые разные стороны. Эти движения объясняются следующим образом. Стилеты соединены скользящими сочленениями, так что их отодвигания друг от друга очень ограничены. Поэтому сильное выдвигание одного стилета может изогнуть весь пучок в противоположную сторону. С другой стороны, длинные стилеты бывают перекручены вдоль продольной оси (рис. 94). Благодаря комбинации этих двух .моментов, концы стилетов и могут быть направляемы в разные стороны.
Дата добавления: 2016-06-29; просмотров: 1555;
Поиск по сайту
Узнать еще
- II. КРОВЬ КАК ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА. ФИЗИОЛОГИЯ ЭРИТРОЦИТОВ И ЛЕЙКОЦИТОВ.
- Автоматическая работа и взаимозависимость клеток организма
- Адсорбция на границе твердое тело – газ
- Адсорбция на границе твердое тело-газ
- Анатомо-топографические принципы кровоснабжения, венозного, лимфатического оттока и иннервации организма человека
- АНТИГЕНЫ ОРГАНИЗМА ЧЕЛОВЕКА
- АНТИГЕНЫ ОРГАНИЗМА ЧЕЛОВЕКА И ЖИВОТНЫХ.
- Антителозависимая клеточная цитотоксичность.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
