Отдел Сумчатые грибы: порядки Гипокреный и Пезизивые. Жизненные циклы Claviceps purpurea, представителей рода Cordyceps. Представители (Morchella conica, Tuber melanosporum).
Порядок гипокрейные – Hypocreales. порядок аскомицетовых грибов класса Sordariomycetes, в составе которого: 7 семейств, 327 родов и 2647 видов. Определить представителя порядка обычно можно по их ярко окрашенному перитециальному плодовому телу или по структуре спорообразовательных систем. Обычно эти грибы имеют жёлтую, оранжевую или красную окраску. Живут как сапротрофы или паразиты растений, грибов и членистоногих. Плодовые тела – перитеции.
Порядки Леотиевые и Пецициевые – Leotiales и Pezizales. Плодовые тела грибов – апотеции. Они мясистой или кожистой консистенции, диаметром 1от 1 мм до 10 см. Аскоспоры освобождаются активно. Исключение - трюфели. У первого порядка сумки открываются порой или трещиной, у второго – крышечкой. Некоторые образуют склероции. Сапротрофы, которые принимают активное участие в разложении растительных остатков, а также паразиты и микоризообразователи. Sclerotinia sclerotiorum, пецица фиолетовая – Peziza violacea, сморчок конический – Morchella conica, сморчковая шапочка - Verpa bohemica, строчек обыкновенный - Gyromitra esculenta. Французский трюфель – Tuber melanosporum.
Жизненный цикл Claviceps purpurea
Образует темные твердые склероции различной формы и размеров в завязи растения-хозяина. Из склероциев после перезимовки развиваются головчатые стромы желтого или красноватого цвета с погруженными в них перитециями. Зимует гриб в почве, куда они попадается при уборке урожая с культурных или дикорастущих злаков, например, на краю поля. Весной склероции прорастают красноватыми головчатыми стромами с перитециями. Далее, аскоспоры заражают злаки во время цветения. Конидии распространяются при трении колосьев друг от друга под действием ветра.
Cordyceps род спорыньёвых грибов, пиреномицеты, паразитирующие на определённых видах насекомых. Кордицепс известен как гриб, обладающий высокими адаптогенными свойствами. Его широкое распространение в мире подтверждает этот факт. Он обитает как в обычных климатических условиях, так и в высокогорных районах на высоте до 6000 метров.
Размножение происходит путём паразитирования на бабочках, мухах и муравьях. Споры, попадающие на волосистую поверхность насекомого, прорастают и внедряются в тело насекомого. Насекомое или личинка бабочки гибнет, и в теле развивается полноценные гифы мицелия кордицепса.
Morchella conica съедобный гриб рода сморчков семейства сморчковых. Плодовое тело — апотеций шляпконожечной формы. «Шляпка» сморчка конического имеет удлинённо-коническую форму; по краю по всей своей окружности плотно прирастает к «ножке». Её высота — 3—9 см, что обычно составляет около 2/3 или 1/2 от высоты ножки, диаметр — 2—4 (до 8) см; окраска — от жёлто-бурой до чёрно-бурая (как правило, коричневого или грязно-коричневого оттенка), иногда серовато-чёрная. Внутри шляпка полая. На поверхности шляпки расположены сильно выступающие складки или рёбра (более тёмные, чем остальная поверхность), которые делят её на более или менее правильно расположенные, вытянутые сверху вниз ячейки. Ячейки различаются по глубине, размеру и форме; выстланы гимением. «Ножка» обычно цилиндрическая, у основания утолщённая или несколько суженная, высотой 2—4,5 см и толщиной 1,5—3 см. Внутри ножка, как и шляпка, полая. Поверхность ножки покрыта продольными бороздками и кажется слегка бархатистой или зернистой. Цвет ножки в зависимости от возраста гриба варьирует от белого до желтоватого или светло-орехового; у молодых грибов он светлее. Подсыхая, ножка сереет и начинает напоминать резину. Мякоть сморчка конического белая или кремовая, тонкая, восковидная, нежная и хрупкая, быстро пересыхающая, внизу ножки водянистая. Выраженным вкусом или запахом не обладает.Споровый порошок светло-охристый или беловато-кремовый, споры (20—24)×(12—14) мкм, эллиптические, гладкие, бесцветные, без капель масла.
Tuber melanosporum. Трюфель черный, гриб рода Трюфель. Плодовое тело (видоизменённый апотеций) подземное, клубневидной, округлой или неправильной формы, 3—9 см в диаметре. Поверхность красновато-коричневого, позже до угольно-чёрного цвета, при надавливании окрашивается в ржавый цвет. Покрыта многочисленными мелкими неровностями с 4—6 гранями. Споры.Мякоть твёрдая, вначале светлая, серая или розовато-коричневая с белым или красноватым мраморным рисунком на срезе, с возрастом темнеет от спор и становится от тёмно-коричневой до чёрно-фиолетовой, прожилки в ней сохраняются. Имеет очень сильный характерный аромат и приятный вкус с горьковатым оттенком.Споровый порошок тёмно-коричневый, споры 35×25 мкм, веретеновидные или овальные, изогнутые.
51. Отдел Базидиальные грибы. а) Общие характерные черты базидиомицетов. б)Строение мицелия, первичный и вторичный мицелий. в) Роль в природе и значение для человека. а) Экология: -среди базидимицетов различают сапротрофов, паразитов (преимущественно растений), симбионтов. Морфология: -Образуют мейоспоры экзогенно на базидиях различного строения (холобазидии/фрагмобазидии или гомобазидии/гетеробазидии); -клеточная стенка хитин-хитин-глюканового типа, в дрожжевой стадии присутствуют маннаны; -многослойная клеточная стенка; -таллом представлен септированным мицелием. Есть дрожжевые формы; -почкование (у дрожжевых форм) происходит с прорыванием материнской клетки и образованием сложного слоистого рубца; -различные септы: простая слоистая с центральной порой может утолщаться (долипоровая), утоньшаться к поре, или иметь равную толщину; -парентесомы – поровые колпачки (мембранные структуры, происходящие из эндоплазматического ретикулюма и закрывающие пору у многих базидиомицетов); -для большинства характерны долипоровые септы с парентосомами (тремелломицеты, гетеробазидиомицеты, гомобазидиомицеты); -для дикариотического мицелия характерны пряжки – клетки, находящиеся у септ мицелия и обеспечивающие параллельное расположение осей деления ядер дикариона вдоль клетки и образование дочерних дикариотических клеток(всегда отсутствуют у ржавчинных, септобазидиальных, гименохетовых, сыроежковых; -есть гомо(10%) и гетероталличные(90%) формы; -дикариотизация несколькими путями: 1)образование анастомозов между двумя гаплоидными мицелиями 2) с образованием специальных половых структур-пикний, содержащих пикноспоры или спермации и воспринимающих гиф;
Размножение:-преобладает дикариотическая фаза; -большинство не имеет дифференцированных половых органов (исключение-ржавчинновые грибы); -половой процесс соматическийб) Мицелий образован тонкими (от 1—2 и до 10—14 мкм, иногда более толстыми) нитями (гифами), которые имеют преимущественно верхушечный рост и густо ветвятся в акропетальном направлении. У базидиальных грибов имеют место два типа мицелия: первичный и вторичный. Каждая клетка первичного мицелия содержит одно гаплоидное ядро. Он обычно недолговечен. У многоих базидиомицетов гаплоидная фаза дрожжеподобная(у многоих головневых грибов). Вторичный (или дикариотический) мицелий образуется после первого этапа полового процесса-плазмогонии. Он долговечен и занимает, порой, очень большое пространство. На нем присутствуют септы.
в) Базидиомицеты в основном сапротрофы, играют важную роль в минерализации органических соединений, в особенности трудноразрушимых (целлюлоза, лигнин). Грибы выделяют свои ферменты в среду и впитывают всей поверхностью продукты распада органики, разрушая при этом гораздо больше вещества чем реально используют. Обитая в почве грибы способны делать ее более плодородной. Уничтожая и минерализуя остатки отмерших организмов, грибы играют очень важную санитарную работу по очистке среды. Плодовые тела многих базидиомицетов съедобны, хотя усвояемость их очень не велика, но они имеют большую пищевую ценность так, как содержат многие незаменимые ферменты и экстрактивные вещества. Отрицательная роль базимицетов тоже очень велика. Грибы трутовики приносят большой вред, лесному хозяйству заражая деревья своей грибницей при этом дерево погибает. Головневые грибы являютя паразитами злаковых. Вызывают порчу продуктов питания.
Отдел базидиальные грибы: особенности полового процесса: утрата типичного полового процесса и переход к соматогамии. Соотношение гаплоидной, диплоидной и дикариотической стадий в цикле развития. Строение плодовых тел. Типы базидий. Строение и типы гименофора.
Бесполое размножение базидиомицетов осуществляется конидиями, но происходит редко. Половое размножение происходит в виде соматогамии, при которой сливаются две вегетативные одноядерные клетки гаплоидного мицелия. У небольшого количества гомоталличных видов могут сливаться клетки одного и того же мицелия. Большинство видов являются гетероталличными, соответственно у них соматогамия происходит только между гифами с противоположными знаками «+» и «-». Половые органы у базидиальных грибов не образуются.
Вначале следует плазмогамия, то есть слияние цитоплазмы клеток. Гаплоидные ядра сближаются, образуя дикарион, но не сливаются между собой. Клетки при этом могут делиться - при этом одновременно митотически делятся и оба ядра дикариона. В результате каждая клетка гифы содержит по дикариону. У большинства видов деление клеток сопровождается образованием боковых выростов - пряжек, что обеспечивает равное деление сестринских ядер и их распределение в дочерние клетки. Стадия дикариона очень длительная, она может продолжаться годами (у некоторых трутовиков десятилетиями). Мицелий при этом растёт, пронизывая субстрат. Дикарионы, кроме того могут образовываться при переносе конидий на мицелий противоположного знака, у некоторых видов (например у головнёвых) могут сливаться базидиоспоры. Органы полового спороношения - базидиоспоры - развиваются экзогенно в особых структурах - базидиях. При этом происходит кариогамия - слияние ядер дикариона, и образуется зигота, которая без периода покоя делится мейотически. Образовавшиеся при этом четыре гаплоидные клетки становятся базидиоспорами, а клетка, от которой они возникли, - базидией. Обычно базидиоспоры располагаются на маленьких и тонких выростах базидии - стеригмах.
Следовательно, в цикле развития базидиальных грибов преобладает дикариотическая фаза. Гаплоидная фаза короткая: базидиоспоры и мицелий, выросший из нее и существующий небольшой период. Диплоидная фаза, как таковая практически отсутствует, т.к. сразу после слияния ядер начинается митотическое деление.
Плодовые тела базидиомицетов различны по форме и консистенции. Они могут быть паутинистыми, рыхлыми, плотно-войлочными, кожистыми, деревянистыми, мягкомясистыми, могут иметь форму пленок, корочек, могут быть копытообразными или состоять из шляпки и ножки. Спороносный слой плодового тела - гимений - располагается у более примитивных видов на верхней стороне плодовых тел, а у более высокоорганизованных - на нижней. Гимений базидиальных грибов состоит из базидий с базидиоспорами и парафиз. У некоторых видов в гимении находятся цистиды - крупные клетки, возвышающиеся над гимениальным слоем. Они защищают гимениальный слой и особенно базидий от давления сверху. Форма цистид для многих видов постоянна и часто служит признаком для их определения. Поверхность плодового тела, несущую гимении, называют гименофором. У низших представителей он гладкий, а у более высокоорганизованных имеет форму зубцов, трубочек, пластинок.
Различают несколько морфологических типов плодовых тел. Распростертые плодовые тела, состоящие из слоя переплетенных гиф, расположенных на субстрате; блюдцевидные или чашевидные плодовые тела с гименофором на внутренней стороне полости; шляповидные плодовые тела - полукруглые, вееровидные, копытовидные, почковидные, прикрепленные боком или зачаточной боковой ножкой с гименофором на нижней стороне плодового тела; шляповидные с центральной ножкой; прямостоячие с гимением, обычно располагающимся в верхней части плодового тела. Между этими морфологическими типами плодовых тел имеются переходные, показывающие постепенное эволюционное развитие плодовых тел от рыхлых, паутинистых, распростертых по субстракту корочек до обособленных, ясно выраженных шляповидных или копытовидных плодовых тел. Параллельно этому шел процесс перемещения гимения с верхней поверхности плодового тела на нижнюю. Расположение гименея на нижней стороне способствует защите спор и их распространению.
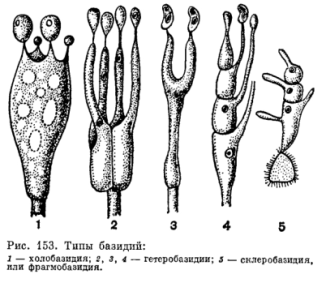
 Базидии делят на типы по морфологии (холобазидии и фрагмобазидии) или развитию (гомобазидии и гетеробазидии). Холобазидия - одноклеточная базидия, не разделенная септами. Фрагмобазидия-разделена поперечными или вертикальными перегородками(обычно на 4 клетки). Гетеробазидия - состоит из гипобазидии и развивающейся из нее эпибазидии, с перегородками или без. Гомобазидия - всегда холобазидия. Не разделяется на гипо- и эпибазидию. Базидия-место кариогамии мейоза и образования базидиоспор, может быть разделена на пробазидию (место кариогамии) и метобазидию (место мейоза). На основании расположения аеретена деления ядер по отношению к продольной оси выделяют два типа холобазидий: хиастобазидии (веретена деления перпендикулярны продольной оси) и стихобазидии(веретена деления параллельны продольной оси). Типичное число спор на базидии равно четырем, но бывают и исключения. Базидиоспоры обычно одноклеточные. Баллистоспоры-базидиоспоры, активно отбрасываемые базидием. Статисмоспора - пассивно освобождающаяся базидиоспора. Акроспоровые базидии - базидиоспоры располагаются на стеригмах на ее вершине на одном уровне. Плевроспоровые базидии - базидиоспоры располагаются на разных уровнях базидия (при наличии перегородок в базидии/замкнутых плодовых телах).
Базидии делят на типы по морфологии (холобазидии и фрагмобазидии) или развитию (гомобазидии и гетеробазидии). Холобазидия - одноклеточная базидия, не разделенная септами. Фрагмобазидия-разделена поперечными или вертикальными перегородками(обычно на 4 клетки). Гетеробазидия - состоит из гипобазидии и развивающейся из нее эпибазидии, с перегородками или без. Гомобазидия - всегда холобазидия. Не разделяется на гипо- и эпибазидию. Базидия-место кариогамии мейоза и образования базидиоспор, может быть разделена на пробазидию (место кариогамии) и метобазидию (место мейоза). На основании расположения аеретена деления ядер по отношению к продольной оси выделяют два типа холобазидий: хиастобазидии (веретена деления перпендикулярны продольной оси) и стихобазидии(веретена деления параллельны продольной оси). Типичное число спор на базидии равно четырем, но бывают и исключения. Базидиоспоры обычно одноклеточные. Баллистоспоры-базидиоспоры, активно отбрасываемые базидием. Статисмоспора - пассивно освобождающаяся базидиоспора. Акроспоровые базидии - базидиоспоры располагаются на стеригмах на ее вершине на одном уровне. Плевроспоровые базидии - базидиоспоры располагаются на разных уровнях базидия (при наличии перегородок в базидии/замкнутых плодовых телах).
Гименофор - поверхность плодового тела, несущая гимений. У низших представителей он гладкий, у более высокоорганизованных бывает трубчатый, пластинчатый, игольчатый, гладкий, лабиринтообразный, сетчатый.
53. Афиллофоидные базидиомицеты:
Гименофор как правило непластинчатый.Плодовые тела в виде подстилки с гименофором, плодовые тела прямостоячие, булавовидной,цилиндрической, кустистой формы, как правило хрящевой или мясистой консистенции. Плодовые тела в виде шляпок с боковой ножкой или без,самой разнообразной формы жесткой консистенции, от кожистой до деревянистой или пробковидной, иногда встречаются плодовые тела со шляпкой и центральной ножкой. Расположение гимения на нижней стороне способствует защите спор и их распространению.
Афиллофороидные макромицеты – индикаторы экологического состояния лесных экосистем
 Гастеромицеты. Мицелий многоклеточный, хорошо развит и разветвлён, пронизывая субстрат. Многие представители способны к образованию мощных тяжей, При помощи тяжей гриб может охватывать значительную территорию, и таким образом они служат для распространения, на тяжах образуются и плодовые тела. Крупные тяжи имеют защитную оболочку из плотно переплетённых толстостенных гиф, преимущественно содержащих отмершие клетки, часто с окрашенными оболочками. Сердцевина тяжа образована более рыхлыми гифами с крупными клетками, выполняет запасающую и проводящую функции. Плодовые тела вначале имеют округлую форму (шаровидную, яйцевидную, грушевидную или цилиндрическую), которая впоследствии может сильно измениться и зависит от механизма освобождения созревших спор. Плодовые тела могут развиваться под землёй или на поверхности, наземные бывают сидячими или имеют настоящую (род Tulostoma) или ложную ножку, образующуюся суженной нижней частью плодового тела. Снаружи плодовое тело покрыто оболочкой — пери́дием, который впоследствии разрывается или образует отверстия для выхода спор. Оболочка может быть однослойной или многослойной, в последнем случае различают наружный экзоперидий и внутренний эндоперидий. При созревании экзоперидий разрывается и остаётся у основания плодового тела наподобие вольвы или растрескивается, обнажая эндоперидий. Эндоперидий служит для защиты глебы и спороносной массы. Он бывает толстым, студенистой консистенции или в виде тонкой мембраны. Раскрывается он отверстиями на вершине плодового тела или неправильными трещинами, может разрываться экваториально, тогда верхняя часть плодового тела исчезает и остаётся чашевидное образование с открыто лежащей споровой массой. Отверстие эндоперидия может образовываться на конце вытянутого «хоботка» — пери́стомы. Внутреннюю мякоть гастеромицетов называют гле́бой. Незрелая глеба состоит из рыхлой однородной трамы, она имеет упруго-мясистую, плотную или студенистую консистенцию. Затем в ней образуются камеры различной формы, выстланные гимениальным слоем с округлыми или неправильной формы холобазидиями. Иногда гимений не образуется и базидии располагаются в камере беспорядочно. При созревании спор глеба разрушается, образуя порошкообразную или слизистую споровую массу. Часть её гиф может сохраняться, образуя капилли́ций — волокнистую массу, разрыхляющую споры. У отдельных видов трама сохраняется и после созревания, внутренность плодового тела у них остаётся плотномясистой. Для некоторых видов гастеромицетов характерно наличие рецепта́кула — плодоносца, выносящего наружу слизистую споровую массу. Компактный вначале рецептакул приобретает ячеистую структуру, быстро увеличивается в размерах и разрывает перидий. Форма его может быть цилиндрической или похожей на шляпконожечный гриб, в виде лопастей или решётчатая. Чаще всего гастеромицеты являются сапрофитами, обитающими в почве и подстилке в лесах и на открытых местностях, в зависимости от условий способны и к микоризообразованию. Среди гастеромицетов нет опасных ядовитых грибов кроме ложнодождевиков. Токсичность их до конца не выяснена, считают, что ложнодождевики могут вызывать нетяжёлые отравления, однако и такие отравления маловероятны из-за неприятного вкуса этих грибов.Съедобны же в молодом возрасте многие дождевики, употребляются в пищу и незрелые плодовые тела некоторых весёлковых,используются в народной медицине.
Гастеромицеты. Мицелий многоклеточный, хорошо развит и разветвлён, пронизывая субстрат. Многие представители способны к образованию мощных тяжей, При помощи тяжей гриб может охватывать значительную территорию, и таким образом они служат для распространения, на тяжах образуются и плодовые тела. Крупные тяжи имеют защитную оболочку из плотно переплетённых толстостенных гиф, преимущественно содержащих отмершие клетки, часто с окрашенными оболочками. Сердцевина тяжа образована более рыхлыми гифами с крупными клетками, выполняет запасающую и проводящую функции. Плодовые тела вначале имеют округлую форму (шаровидную, яйцевидную, грушевидную или цилиндрическую), которая впоследствии может сильно измениться и зависит от механизма освобождения созревших спор. Плодовые тела могут развиваться под землёй или на поверхности, наземные бывают сидячими или имеют настоящую (род Tulostoma) или ложную ножку, образующуюся суженной нижней частью плодового тела. Снаружи плодовое тело покрыто оболочкой — пери́дием, который впоследствии разрывается или образует отверстия для выхода спор. Оболочка может быть однослойной или многослойной, в последнем случае различают наружный экзоперидий и внутренний эндоперидий. При созревании экзоперидий разрывается и остаётся у основания плодового тела наподобие вольвы или растрескивается, обнажая эндоперидий. Эндоперидий служит для защиты глебы и спороносной массы. Он бывает толстым, студенистой консистенции или в виде тонкой мембраны. Раскрывается он отверстиями на вершине плодового тела или неправильными трещинами, может разрываться экваториально, тогда верхняя часть плодового тела исчезает и остаётся чашевидное образование с открыто лежащей споровой массой. Отверстие эндоперидия может образовываться на конце вытянутого «хоботка» — пери́стомы. Внутреннюю мякоть гастеромицетов называют гле́бой. Незрелая глеба состоит из рыхлой однородной трамы, она имеет упруго-мясистую, плотную или студенистую консистенцию. Затем в ней образуются камеры различной формы, выстланные гимениальным слоем с округлыми или неправильной формы холобазидиями. Иногда гимений не образуется и базидии располагаются в камере беспорядочно. При созревании спор глеба разрушается, образуя порошкообразную или слизистую споровую массу. Часть её гиф может сохраняться, образуя капилли́ций — волокнистую массу, разрыхляющую споры. У отдельных видов трама сохраняется и после созревания, внутренность плодового тела у них остаётся плотномясистой. Для некоторых видов гастеромицетов характерно наличие рецепта́кула — плодоносца, выносящего наружу слизистую споровую массу. Компактный вначале рецептакул приобретает ячеистую структуру, быстро увеличивается в размерах и разрывает перидий. Форма его может быть цилиндрической или похожей на шляпконожечный гриб, в виде лопастей или решётчатая. Чаще всего гастеромицеты являются сапрофитами, обитающими в почве и подстилке в лесах и на открытых местностях, в зависимости от условий способны и к микоризообразованию. Среди гастеромицетов нет опасных ядовитых грибов кроме ложнодождевиков. Токсичность их до конца не выяснена, считают, что ложнодождевики могут вызывать нетяжёлые отравления, однако и такие отравления маловероятны из-за неприятного вкуса этих грибов.Съедобны же в молодом возрасте многие дождевики, употребляются в пищу и незрелые плодовые тела некоторых весёлковых,используются в народной медицине.
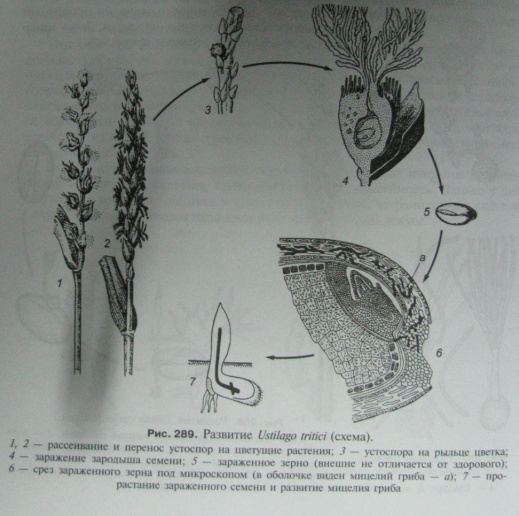
54. Головневые грибы. Морфология:Нежный, септированный мицелий головневых проникает внутрь клеток или проходит по межклетным пространствам. Микроскопически малые споры в большинстве случаев развиваются только в известных частях заболевшего растения; Размножение: Образование спор у большинства головневых влечет за собой различные патологические изменения тканей заболевшего растения (иногда гипертрофии и своеобразные выростки), а потом и полную их дезорганизацию и разрушение. Из споры (хламидоспоры) в подходящей питательной среде развивается короткий росток (промицелий). Росток этот разгораживается тремя или четырьмя поперечными перегородками на отдельные клеточки, отшнуровывающие повторно сбоку овальные конидии ("споридии"). Отпавшие конидии размножаются почкованием наподобие дрожжей. По мере того как запас питательного материала в окружающей среде истощается, конидии перестают почковаться и вырастают в нити. Когда нить встретит молодой, недавно вышедший из семени росток, она прободает его эпидерм, входит внутрь и направляется к точке роста, к месту, где впоследствии сформируется соцветие. Таким образом происходит заражение. Нити мицелия (гифы) обильно ветвятся внутри завязи и под конец совершенно разрушают ее ткань. Внутри нитей образуются споры. Когда споры созреют, оболочка гиф разрушается и споры высыпаются. каждого вида Головневые грибы хламидоспоры могут быть только в определенных органах растения-хозяина. Половой процесс Головневые грибы состоит в копуляции двух базидиоспор или отпочковавшихся от них клеток , причем содержимое из одной споры переходит в другую, но ядра не сливаются. Из образовавшейся диплоидной клетки развивается грибница с двухъядерными клетками, способная заразить растение.
Таким образом, цикл развития состоит в чередовании гаплоидной (базидии и базидиоспоры) и диплоидной (грибница и хламидоспоры) фаз. Основное значение в жизни Головневые грибы имеет диплоидная фаза, которая начинается копуляцией базидиоспор и заканчивается редукционным делением ядра в хламидоспоре.
Значение для природы и человека:паразиты высших растений, вызывающие у них заболевания, известные под назв. головни (рис. 1). Особенно большой вред наносят Головневые грибы хлебным злакам.
Твердая головня (Tiletia caries) проходит свой цикл развития за один вегетационный период. На почве весной телиоспоры прорастают в фрагмобазидии с четырьмя стеригмами, несущими базидиоспоры. Эти базидиоспоры попарно копулируют. В таком состоянии они напоминают букву Н. При прорастании зерна гриб проникает в растение (проростковая инфекция), образуя дикарионный мицелий. Достигнув конуса нарастания, мицелий заполняет развивающиеся зерновки. Колос пораженного растения не поникает под тяжестью зерна, потому что зерновки, наполненные телиоспорами легкие.
Ржавчиновые грибы.
Экология: В условиях умеренного климата болезнь на ржи появляется как правило в фазе колошения. Урединиоспоры прорастают и внедряются в лист при температуре от 5 до 25. C (лучшая температура 18 . 22.C) при условии наличия капельно-жидкой влаги (росы). При благоприятных условиях уредогенерация развивается за 7 дней, то есть, за сезон возможно развитие нескольких генераций. Распространение урединиоспор осуществляется ветром на небольшие или дальние (сотни километров) дистанции. Озимая рожь предоставляет грибу возможность зимовать в стадии уредомицелия в листьях ржи, если заражение произошло с осени. Таким образом отсутствие промежуточного хозяина не влияет на распространение болезни в различных географических зонах.
Морфология: Мицелий эндофитный, межклетный, с внедряющимися в полость клеток гаусториями, через которые гриб поглощает питательные вещества. В клетках мицелия и особенно в спорах большинства Ржавчинные грибы содержится большое количество масла в виде капель, окрашенных липохромом в оранжевый или ржавый цвет (отсюда название "Ржавчинные грибы"). У большинства Ржавчинные грибы мицелий сосредоточен в месте проникновения инфекции: обычно после образования спороношений гриб вызывает гибель ткани растения-хозяина, после чего отмирает и сам.инфекции: обычно после образования спороношений гриб вызывает гибель ткани растения-хозяина, после чего отмирает и сам. Некоторым Ржавчинные грибы свойствен диффузный мицелий, который пронизывает ткань всего растения; он часто бывает многолетним и нередко вызывает значительные изменения в анатомическом строении пораженных органов (опухоли, "ведьмины метлы").
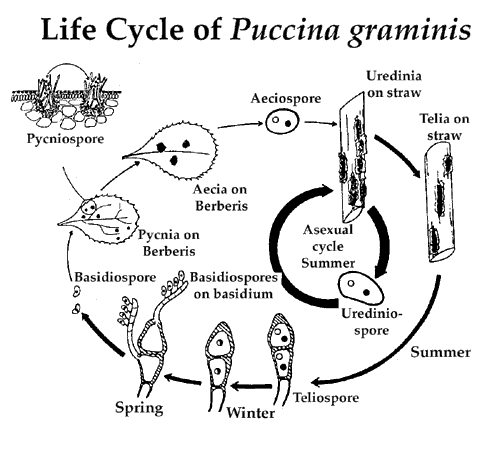 Размножение: перезимовавшие теллоспоры прорастают с образованием базидий с базидиоспорами (в ней перед образованием базидиев происходит кариогамия, образуется метабазидия, в которой происходит мейоз). Базидии –фрагмобазидии, разделенные поперечными перегородками на 4 клетки, образующие по одной базидиоспоре, в которой находится гаплоидное ядро. После созревания разбрасываются, попадают на листья барбариса и формируют весеннюю фазу развития: ростковая трубка проникает в лист, мицелий распространяется по межклетникам, запуская в клетки лаусториии. Гифы мицелия гаплоидны.На этом этапе в тканях гриба закладывается 2 типа спороношения: на верхней стороне листа образуются клубочки гиф, превращающиеся в пикнии, в которых образуются пикниоспоры, выполняющие функции спермациев, при их созревании из отверстия пикния высовываются воспринимающие гифы и сладкая жидкость, привлекающая насекомых –переносчиков пикноспор (т.к. гриб гетероталломен, для дикариотизации нуэны разные особи); после дикариотизации на нижней стороне листа развивается следующий тип спороношения – эцидии. Они закладываются еще на гаплоидной стадии, но оформляются только после дикариотизации; эцидии имеют вид чашечек, окруженных перидием, образующегося из срастающихся цепочек двухъядерных клеток. Цепочки развиваются из краевых базальных клеток эцидия (оба ядра базальной клетки делятся одновременно и ее верхняя часть отделяется в виде двухъядерной материнской клетки эцидиоспоры, процесс повторяется многократно. Каждая материнская клетка эцидиоспоры делится, образуя 2 двухъядерные клетки неравного размера (верхняя более крупная с толстостенной оболочкой превращается в эцидиоспору. Нижняя тонкостенная быстро разрушается, верхние эцидиоспоры цепочек страстаются, образуя крышку эция; чешевидные эции, одетые перидием, называются эцидиями; нарастающие цепочки эциоспор прорывают эпидермис листа и крышку перидия. По мере созревания, эциоспоры отцепляются и переносятся по воздуху. Эциоспоры одноклеточные, не могут заражать барбарис. Они должны попасть на вегетативные органы злаков, где ростковыми трубками они прорастают с образованием локального дикариотического мицелия с гаусториями. На этом мицелии образуются урединиоспоры. Образуются в массе под эпидермисом листа или стебля, а затем прорывают его и выступают в виде продолговатых пустулурединиев, уредиоспоры могут повторно поражать злаки, по мере истощения субстрата на том же мицелии к концу лета образуются телиоспоры (двуклеточные, расположены на ножках в телиях с одной ростковой парой в каждой клетке, покрывающая споры для зимовки) сначала телиоспоры содержат дикарион, но затем происходит кариогамия с образованием диплоидного ядра. Весной каждая клетка проростает четырехклеточной базидией, куда переходит диплоидное ядро, делящееся в базидии митозом. Гаплоидные ядра переходят в базидиоспоры, которые снова заражают барбарис.
Размножение: перезимовавшие теллоспоры прорастают с образованием базидий с базидиоспорами (в ней перед образованием базидиев происходит кариогамия, образуется метабазидия, в которой происходит мейоз). Базидии –фрагмобазидии, разделенные поперечными перегородками на 4 клетки, образующие по одной базидиоспоре, в которой находится гаплоидное ядро. После созревания разбрасываются, попадают на листья барбариса и формируют весеннюю фазу развития: ростковая трубка проникает в лист, мицелий распространяется по межклетникам, запуская в клетки лаусториии. Гифы мицелия гаплоидны.На этом этапе в тканях гриба закладывается 2 типа спороношения: на верхней стороне листа образуются клубочки гиф, превращающиеся в пикнии, в которых образуются пикниоспоры, выполняющие функции спермациев, при их созревании из отверстия пикния высовываются воспринимающие гифы и сладкая жидкость, привлекающая насекомых –переносчиков пикноспор (т.к. гриб гетероталломен, для дикариотизации нуэны разные особи); после дикариотизации на нижней стороне листа развивается следующий тип спороношения – эцидии. Они закладываются еще на гаплоидной стадии, но оформляются только после дикариотизации; эцидии имеют вид чашечек, окруженных перидием, образующегося из срастающихся цепочек двухъядерных клеток. Цепочки развиваются из краевых базальных клеток эцидия (оба ядра базальной клетки делятся одновременно и ее верхняя часть отделяется в виде двухъядерной материнской клетки эцидиоспоры, процесс повторяется многократно. Каждая материнская клетка эцидиоспоры делится, образуя 2 двухъядерные клетки неравного размера (верхняя более крупная с толстостенной оболочкой превращается в эцидиоспору. Нижняя тонкостенная быстро разрушается, верхние эцидиоспоры цепочек страстаются, образуя крышку эция; чешевидные эции, одетые перидием, называются эцидиями; нарастающие цепочки эциоспор прорывают эпидермис листа и крышку перидия. По мере созревания, эциоспоры отцепляются и переносятся по воздуху. Эциоспоры одноклеточные, не могут заражать барбарис. Они должны попасть на вегетативные органы злаков, где ростковыми трубками они прорастают с образованием локального дикариотического мицелия с гаусториями. На этом мицелии образуются урединиоспоры. Образуются в массе под эпидермисом листа или стебля, а затем прорывают его и выступают в виде продолговатых пустулурединиев, уредиоспоры могут повторно поражать злаки, по мере истощения субстрата на том же мицелии к концу лета образуются телиоспоры (двуклеточные, расположены на ножках в телиях с одной ростковой парой в каждой клетке, покрывающая споры для зимовки) сначала телиоспоры содержат дикарион, но затем происходит кариогамия с образованием диплоидного ядра. Весной каждая клетка проростает четырехклеточной базидией, куда переходит диплоидное ядро, делящееся в базидии митозом. Гаплоидные ядра переходят в базидиоспоры, которые снова заражают барбарис.
Значение: Стеблевая ржавчина одно из наиболее опасных заболеваний ржи. Болезнь увеличивает водообмен, уменьшает уровень метаболитов доступных для формирования зерна, вызывает ломкость стеблей, что приводит к уменьшению числа зерен в колосе и их веса. Снижение урожая достигает 10-20%, а в условиях эпифитотии более 35%
Дата добавления: 2016-05-30; просмотров: 5395;
Поиск по сайту
Узнать еще
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- II. Степень насыщенности, определяемая природой связи между атомами углерода.
- II. Установление сегуната Токугава. Социально-экономическое развитие страны: циклы подъема и упадка
- II.4. Природа резус-несовместимости при гемотрансфузии
- III Отделение Собственной Его Императорского Величества канцелярии
- III. Ветви третьего отдела.
- III. Функции отдела по делам ГОЧС и ВМР

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
