Дифференцировка зародышевых листков
| Зародышевый листок | Производные во взрослом организме | |||
| ЭКТОДЕРМА | Покровная эктодерма | эпидермис кожи и его производные (волосы, ногти, потовые и сальные железы); молочные железы; эпителиальная выстилка органов полости рта; энамелобласты (секреция органического матрикса эмали зуба); слюнные железы; хрусталик глаза; аденогипофиз; выстилка наружного, среднего и внутреннего уха; эпителий анального канала | ||
| Нейроэктодерма | Нервная трубка | центральная нервная система (все нейроны и глия); сетчатка; нейрогипофиз; эпифиз | ||
| Нервный гребень | периферическая нервная система и другие структуры (таблица | |||
| МЕЗОДЕРМА | Параксиальная (СОМИТЫ) | Дерматом | Дерма кожи | |
| Миотом | Скелетная мускулатура туловища, конечностей, головы и шеи; глазные мышцы; мышечное тело языка. | |||
| Склеротом | Позвонки, рёбра, лопатки | |||
| Промежуточная (НЕФРОТОМ) | Эпителий почки и органов репродуктивной системы | |||
| Латеральная (СПЛАНХНОТОМ) | Выстилка серозных оболочек полостей тела (брюшина, перикард, плевра) Миоэпикардиальная пластика (миокард, эпикард) Корковое вещество надпочечников | |||
| ЭНТОДЕРМА | эпителиальная выстилка желудочно-кишечного тракта, печень, поджелудочная железа, желчный пузырь; эпителиальная выстилка дыхательной системы (трахея, бронхи, лёгкие), строма тимуса, эндокриноциты щитовидной и паращитовидной железы, эпителий мочевого пузыря, женской уретры и большей части мужской уретры. | |||
| МЕЗЕНХИМА | Кровь, лимфа, соединительная ткань, гладкая мышечная ткань, кровеносные сосуды, эндокард | |||
Таблица 5
| ПРОИЗВОДНЫЕ НЕРВНОГО ГРЕБНЯ |
| Чувствительные нейроны спинальных узлов и узлов черепных нервов Нейроны симпатической и парасимпатической нервной системы Нейролеммоциты (шванновские клетки) и глиоциты ганглия Мягкая и паутинная оболочки мозга; Меланоциты Клетки каротидного тельца Парафолликулярные клетки щитовидной железы Клетки диффузной эндокринной системы желудочно-кишечного и дыхательного трактов (APUD-серия) Хромаффиноциты мозгового вещества надпочечников Хрящи, кости, мышцы и соединительные ткани лица (эктомезенхима) Верхнечелюстной и челюстной отростки; Подъязычные дуги и 3-я глоточная дуга; Одонтобласты → дентин зуба Задний эпителий роговицы Расширяющая и суживающая мышцы зрачка Ресничная мышца |
Embryology is a science about the embryonic development of the organism.
The embryonic development, or embryogenesis, is a period from the moment of fertilization to the birth (in live-bearing animals).
The process of embryogenesis may be divided into several stages:
· fertilization is a series of events leading to fusion of male and female gametes with formation of unicellular new organism, the zygote;
· cleavage, which consists of a series of rapid mitotic divisions which result in in formation of multicellular embryo, the blastula;
· gastrulation is a complex series of morphogenetic movements of cell groups leading to the establishment of the primary germ layers that giving rise to the embryo, composing of three germ layers, the gastrula;
· neurulation and notogenesis,theformation of the nerve tube and other axial primordia;
· histogenesis and organogenesis,each of the three germ layers gives rise to a number of specific tissues and organs;
· systemogenesis, the organ systems formation..
Gametogenesis,the production of specialized male and female germ cells or gametes precedes fertilization.
GAMETOGENESIS. REPRODUCTIVE CELLS
Gametogenesis is the production of specialized germ cells, or gametes. During the gametes’ maturation the number of chromosomes is reduced by half to the haploid number.
Primordial germ cells, lso known as PGCs,precursor germ cells or gonocytes orgonoblasts,in human embryos appear in the wall of the yolk sac at the end of the third week of the prenatal life. These cells are indifferent: they are not divided onto the male and the female cells. They migrate by ameboid movement from the yolk sac toward through tthe developing gonads (the paired gonadal ridges on the medial surface of the mesonephros), divide repeatedly on their migratory route, and differentiate there into spermatogonia or oogonia (in humans, sexual differentiation starts approximately 6 weeks post conception). Germ cells produce gametes and are the only cells that can undergo meiosis as well as mitosis.
SPERMATOGENESIS
Male reproductive cells, the spermatozoa, are produced in seminiferous tubules of the testis beginning with the puberty age and continue throughout adult life. The process of differentiation of the male germ cells is called spermatogenesis and can be divided into 4 phases (Fig.2):
· proliferation phase
· growth phase
· maturation phase
· formation,orspermiogenesis
The phase of proliferation.During this phase spermatogonia divide by mitoses, producing successive generations of cells that finally give rise to primary spermatocytes.Spermatogonia consist of two types: type A spermatogonia, and type B spermatogonia (fig.1).
· Type A dark spermatogonia are true stem cells, steady to harmful factors action, rarely dividing cell. At sexual maturity, this cell undergoes a series of mitoses, and the newly formed cells can follow one of 2 paths: they can continue, as undifferentiated stem cells (type A pale spermatogonia), or they can differentiate during progressive mitotic cycles to become type B spermatogonia.
· Type B spermatogonia are committed cells, they prepare to the first meiotic division, enter the phase of growth and give rise to primary spermatocytes.
The phase of growth. During this phase primary spermatocytes grow (their size 4 times more as spermatogonia), and enter a prolonged prophase of the first meiotic division (lasts about 22 days), followed by rapid completion of meiosis I and formation of secondary spermatocytes;
The phase of maturation. Secondary spermatocytes immediately enter the secondmeiotic division, to form spermatids, which contain the haploid number of chromosomes;
The phase of formation,orspermiogenesis.During this phase the spermatids go through an elaborate process of cytodifferentiation, producing small, highly motile spermatozoa.
Spermiogenesisis a complex process of differentiation that includes (Fig.3):
· formation of the acrosome;
· condensation and elongation of the nucleus;
· flagellum’ development;
· the loss of much of the cytoplasm.
The acrosome is a special organelle, originated from the Golgi complex. It is a membrane-limited vesicle (the acrosomal cap), which cover the anterior half of the condensing nucleus. The acrosome contains several hydrolytic enzymes, such as hyaluronidase, penetrase, acrosin (trypsin-like e  nzyme), acid phosphatase, and proteases. The acrosome thus serves as a specialized type of lysosomes. These enzymes are known to dissociate follicular cells of the corona radiata and to digest the zona pellucida, structures that surround recently ovulated eggs.
nzyme), acid phosphatase, and proteases. The acrosome thus serves as a specialized type of lysosomes. These enzymes are known to dissociate follicular cells of the corona radiata and to digest the zona pellucida, structures that surround recently ovulated eggs.
Fig.3. Top: The principal changes occurring in spermatids during spermiogenesis.
Bottom: The structure of a spermatozoon.
During spermiogenesis the centrioles of the spermatid migrate to a position near the cell surface and opposite the location of the forming acrosome. One of the centrioles (distal) gives rise to microtubules of theaxonemeoftheflagellum. The axoneme is surrounded by dense skeleton (coarse fibrils, rib-like fibrils and columns). The mitochondria aggregate in a helical manner around the proximal part of the flagellum axoneme, forming a thickened region known as the middle piece, the region where the movements of the spermatozoa are generated.
The reduced volume of the nucleus permits the sperm greater mobility and may protect the genome from the damage while it transit to the egg cell.
Residual cytoplasm is shed and excess cytoplasm is phagocytosed by Sertoli cells. Cytoplasmic bridges break down and the spermatozoa are released into the lumen of the seminiferous tubule.
The time required for a spermatogonium to become a mature spermatozoon is approximately 64 days.
During division of the spermatogonia, the resulting cells do not separate completely but remain attached by cytoplasmic bridges forming non-cellular structure called the syncitium. The intercellular bridges provide communication between every primary and secondary spermatocytes and spermatid derived from a single spermatogonium. By permitting the interchange of information from cell to cell, these bridges play an important role in coordinating the sequence of events in spermatogenesis. When the process of spermatogenesis is completed, the sloughing of the cytoplasm and cytoplasmic bridges as residual bodies leads to a separation of the spermatids.
The mature spermatozoon has a lash-like appearance and consists of a head, a neck, and a tail with middle, principal and terminal pieces (Fig.4). The head, which, for most mammals, has a flattened, oval shape, contains the condensed nucleus and is capped by an acrosome filled with hydrolytic enzymes. The midpiece contains large helical mitochondria that generate the energy. The tail contains microtubules.
OOGENESIS
.The process of oogenesis occurs in the ovary. The oogenesis includes 3 phases (Fig.4):
· prolif
· growth
· maturation
The phase of proliferation. Once primordial germ cells have arrived in the gonad of a genetic female, they differentiate into oogonia. Oogonia divide by mitosis only during embrional life. Beginning in the 3-rd fetal month, some oogonia enter the prophase of the 1st meiotic division and become much larger primary oocytes. In the human, this process is completed by the end of the seventh month of gestation. During this time, many primary oocytes are lost as a result of a degenerative process called atresia.So oocytes are formed during intrauterine life, and their number does not increase after birth.
The phase of growth. This phase lasts a very long time (from 12 up to 50 years) and subdivided onto small and large growth.
Small growth period. At birth all egg cells in the ovary are primary oocytes. Their total number varies from 700,000 to 2 million.
The primary oocytes become surrounded by a layer of flattened follicular cells and become known as primordial follicles. The process of encapsulation of the primary oocyte blocks the first meiotic division in the prophase. Primary oocytes do not finish their 1st meiotic division but remain in the diplotene stage until puberty.
Large growth period. With the onset of puberty a number of primordial follicles begin to grow and mature with each ovarian cycle under influence of follicle-stimulating hormone (FSH). Under normal conditions, only one of these follicles reaches full maturity, the others degenerate and become atretic. Only approximately 400,000 primary oocytes are present by the beginning of puberty, and fewer than 500 will be ovulated in the reproductive lifetime of the individual.
The phase of maturation. The first meiotic division is completed just before ovulation. The chromosomes are equally divided between the daughter cells, but one cell, the secondary oocyte, retains almost all of the cytoplasm. The other becomes the first polar body, a very small cell containing the nucleus and a minimal amount of cytoplasm. Immediately after expulsion of the first polar body the secondary oocyte starts the 2nd meiotic division, which stops in metaphase. The 2nd meiotic (maturation) division will be completed only if the oocyte is fertilized. The secondary oocyte, in turn, gives rise to a mature oocyte plus another polar body. Hence, a primary oocyte develops into one mature oocyte and three polar bodies.
Зрелая яйцеклетка млекопитающих – крупная, в сравнении со сперматозоидом, неподвижная клетка. Особенность строения яйцеклетки – отсутствие центриолей и наличие характерных трофических включений – желточных гранул. По периферии яйцеклетки, под клеточной мембраной, равномерно распределены кортикальные гранулы, содержащие ферменты (гидролазы), необходимые для формирования оболочки оплодотворения после проникновения сперматозоида.
При овуляции яйцеклетка окружена (рис.6):
• прозрачной оболочкой (zona pellucida),содержащей густую сеть тонких нитей, состоящих из гликопротеинов (фракции ZP1, ZP2, ZP3);
фолликулярными клетками, образующими лучистый венец
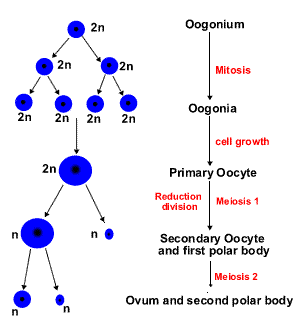
The phase of formation in oogenesis is lacking.
Table 1
| Comparison of spermatogenesis and oogenesis | ||
| Similarities | Differences | |
| Spermatogenesis | Oogenesis | |
| Both processes begin with mitosis (phase of proliferation) Both processes have meiosis | · Meiosis I occurs only at puberty · Fertility is life long but reduces · Different timing of maturation: spermatogenic meiosis is rapid and uninterrupted. · Spermatogenesis has equivalent meiotic divisions: spermatogonium gives rise to 4 spermatids | · Meiosis I begins in embryogenesis (the 5th month) · Fertility is limited · Oogenic meiosis is interrupted at one or more stages (for a long time, up to 50 years) · Oogenic meiosis is asymmetrical: only one egg is formed together with 3 polar bodies. · The phase of formation is lacking. |
Ovary Structure
Oogenesis results in a large non-motile ovum(Fig.5). This cell is spherical and many times larger than a spermatozoon. Ovum has no centrioles, so this cell is not capable to divide itself. The cytoplasm of the ovum is packed with food reserve. Membrane-bound cortical granules accumulate in the cytoplasm, just beneath the plasma membrane. Cortical granules contain enzymes releasing a cortical reaction.
Fig.5. Human egg cell: 1) nucleus; 2) egg cell microvilli; 3) follicular cells; 4) follicular cells processes; 5) cortical granules; 6) yolk inclusions; 7) zona pellucida; 8) receptors.
In matu  re follicles the oocyte is enveloped by the zona pellucida, a thick coat, composed of at least 3 different glycoproteins (ZP1, ZP2, ZP3). ZP3 is the ligand in the zona pellucida which binds with the acrosome of the sperm in the acrosome reaction.
re follicles the oocyte is enveloped by the zona pellucida, a thick coat, composed of at least 3 different glycoproteins (ZP1, ZP2, ZP3). ZP3 is the ligand in the zona pellucida which binds with the acrosome of the sperm in the acrosome reaction.
The oocyte with the zona pellucida is surrounded by several layers of follicular cells. Long apical processes of follicular cells and microvilli of the oocyte penetrate the zona pellucida and make contact with one another via gap junctions. The follicular cells of the first layer around the ovum, that are, therefore, in close contact with the zona pellucida form the corona radiata, which accompanies the ovum when it leaves the ovary in the process of ovulation. The corona radiata is still present when the spermatozoon fertilizes the ovum; it is retained for some time during the passage of the ovum through the oviduct.
FERTILIZATION
Fertilization (also known as conception), the process by which male and female gametes fuse, occurs normally in the in the ampulla of the uterine tube.
During ovulation the secondary oocyte, together with surrounding the zona pellucida and corona radiata cells, is discharged from the ovary and is carried into the tube by sweeping movements of the fimbriae and by motion of cilia on the epithelial lining.
Human ejaculate contains 3x108 spermatozoa. These cells remain viable in the female reproductive tract for approximately 24 hours. Only 300-500 spermatozoa reach the site of fertilization.
Fertilization is more a chain of events than a single, isolated phenomenon and may be subdivided into 3 stages:
· distant interaction;
· contact interaction;
· penetration of the sperm cell into the oocyte and egg activation;
· fusion of male and female pronuclei, and formation of a zygote.
The process of fertilization lasts 12-24hours.
The distant interaction is based on phenomenon of chemotaxis: it was demonstrated that the surface membrane of human sperms possess receptors for specific chemoattractant substances (such as formyl Met-Leu-Phe) which have the ability to induce migration of sperm. Contraction of the musculature of the uterine tube promote this ascent.
Freshly ejaculated sperm are unable to fertilize. Sperm cells must undergo capacitation.
Capacitationoccurring in the female reproductive tract, is a complex process, which renders the sperm cell capable for specific interaction with the oocyte. This process takes place under influence of mucous secretion of uterine tube epithelial cells, pH in the female reproductive tract etc. This final step in the maturation of spermatozoa is biochemical event. It associated with removal of adherent seminal plasma glycoproteins, reorganization of plasma membrane lipids and proteins, rendering it more fusigenic. The result is a more fluid membrane with an increased permeability to Ca2+. An influx of Ca2+ produces increased intracellular cAMP levels and thus, an increase in motility. Hyperactivation is also part of capacitation and is the result of the increased Ca2+ levels.
Capacitation lasts approximately 7 hours. Spermatozoa used for in vitro fertilization are artificially capacitated.
The contact interaction includes:
· attachment and binding. The sperm binds to the zona pellucidaof the egg. A special complementary molecule on the surface of the sperm head then binds to ZP3 glycoproteins in the zona pellucida. Binding of sperm to the zona pellucida is a receptor-ligand interaction with a high degree of species specificity: only sperm of same species may enter.
· the acrosomal reaction.This binding triggers the acrosomal reaction: the acrosome vesicle membrane fuses with the sperm plasma membrane at multiple sites, releasing acrosomal enzymes, such as hyaluronidase and acrosin to degrade the zona pellucida and, in that way, facilitate penetration through it. After the acrosome reaction, it is believed that the sperm remains bound to the zona pellucida through exposed ZP2 receptors.
Penetration of spermatozoon and egg activation and the cortical reaction. Upon binding of a sperm cell, the egg rapidly undergoes a number of metabolic and physical changes that collectively are called egg activation. Prominent effects include a rise in the intracellular concentration of calcium, completion of the second meiotic division and the cortical reaction. Once the sperm cells find their way past the zona pellucida (owing to 2 factors: motility and zona-digesting enzymes), the cortical reaction occurs: cortical granules inside the ovum fuse with the plasma membrane of the cell, causing enzymes (including several proteases) inside these granules to be expelled to the zona pellucida. This in turn causes the glycoproteins in the zona pellucida to cross-link with each other, making the whole matrix hard and impermeable to sperm (sperm receptors are destroyed also). So, the zona pellucida transforms into the fertilization membrane (or the zona reaction) around a fluid-filled perivitelline space. This membrane serves as a barrier and prevents completely further penetration of other sperms (block to polyspermy).
Дата добавления: 2020-05-20; просмотров: 1019;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
