Структура клеточных стенок прокариот
Большинство прокариот имеют на поверхности плазматической мембраны жесткую, многослойную клеточную стенку, которая выполняет функцию механической защиты клетки, а также придает ей форму. Клеточная стенка может быть отделена от мембраны периплазматическим пространством, в котором находятся ферменты и белки, связывающие питательные вещества. В отличие от плазматической мембраны, клеточная стенка проницаема для солей и низкомолекулярных соединений. Основу клеточных стенок большинства прокариот составляет пептидогликан муреин. Однако состав и структура клеточных стенок двух самых представительных отделов прокариот (Gracilicutes и Firmicutes) сильно различается, что положено в основу дифференциального метода окраски бактерий по Граму.
Клеточная стенка грамположительных бактерий. Практически целиком состоит из муреина. Этот гетерополимер содержит чередующиеся остатки N-ацетилглюкозамина и N-ацетилмурамовой кислоты, связанные между собой b(1→4)-гликозидными связями. Молекула муреина уникальна по своему строению, поскольку содержит D-аминокислоты, диаминопимелиновую кислоту и g-пептидную связь (рис. 5.4).
Необычность строения муреина выражается, в первую очередь, в особенностях тетрапептида в составе N-ацетилмурамовой кислоты. Первую позицию в нем занимает L-аланин, вторую — D-изоглутаминовая кислота (либо какая-нибудь минорная аминокислота). Изоглутаминовая кислота формирует пептидную связь с L-лизином с помощью своей g-кар-
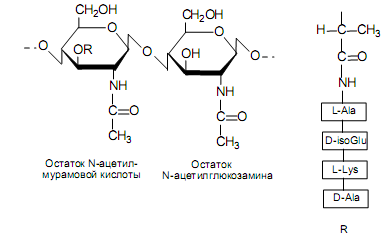
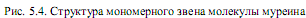
боксильной группы (g-пептидная связь), а не a-карбоксильной группы, как обычно. Третью позицию в тетрапептиде занимает L-лизин, который имеет боковую аминогруппу, хотя он может быть замещен на другие кислоты с боковой аминогруппой (диаминопимелиновую или диаминомасляную) (рис. 5.5). Последнюю позицию в тетрапептиде всегда занимает D-аланин. D-Аминокислоты в составе муреина придают ему повышенную устойчивость к протеолитическим ферментам.
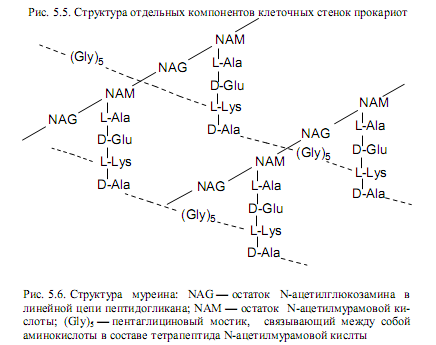
Линейные цепи пептидогликана, в которых чередуются дисахаридпептидные звенья (рис. 5.4), составляют основу муреина. Его структуру усложняют короткие пептиды, поперечно сшивающие полисахаридные цепи. Благодаря множеству поперечных сшивок возникает одна огромная мешковидная макромолекула, которую называют муреиновым каркасом. Поперечные сшивки чаще всего представлены пентаглициновыми пептидами (мостиками), но в их состав могут входить и другие аминокислоты: L-аланин, L-треонин или L-серин. При формировании сшивок карбоксильная группа одного концевого остатка глицина в пентаглициновом мостике образует пептидную связь с боковой аминогруппой лизина в составе тетрапептида N-ацетилмурамовой кислоты какой-либо цепи, а свободная аминогруппа другого концевого остатка в пентаглициновом мостике образует пептидную связь со свободной карбоксильной группой концевого остатка D-аланина в составе N-ацетилмурамовой кислоты той же или другой муреиновой цепи (рис. 5.6). Таким образом, один тетрапептид может участвовать в образовании сразу двух сшивок между цепями с помощью пентаглициновых мостиков.
Сформированная в результате сложная муреиновая сеть упрочняется тейхоевыми кислотами, которые покрывают слои пептидогликана или инкрустируют их. Тейхоевые кислоты представляют собой цепочки из молекул глицерола или пятиатомного спирта рибитола, связанные между
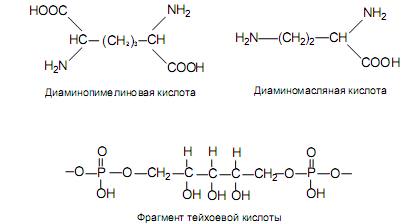
собой фосфодиэфирными мостиками (рис. 5.5). В одной молекуле тейхоевой кислоты встречается около 30 остатков спиртовых молекул. В эту цепочку могут быть включены остатки молекул аминокислот или сахаров. Гидроксильные группы спиртов в составе тейхоевых кислот используются для связывания пептидогликановых слоев друг с другом и делают муреиновый каркас более прочным.
Всего в составе клеточной стенки грамположительных бактерий насчитывается около 40 слоев муреина, и он составляет 30—70% сухой массы клеточной стенки. Стенка сохраняет прочность и форму клетки даже в том случае, когда клеточное содержимое ликвидируется, например растворяется кислотой.
Клеточная стенка грамотрицательных бактерий. У грамотрицательных бактерий клеточная стенка организована более сложно, чем у грамположительных. Ее основными отличительными особенностями являются следующие: муреиновая сеть однослойная и составляет только 5— 10% сухой массы клеточной стенки; в стенке отсутствуют тейхоевые кислоты; на поверхности пептидогликанового слоя есть богатая липидами наружная мембрана (рис. 5.7). По структуре муреин грамотрицательных бактерий отличается от такового грамположительных низким содержанием поперечных сшивок и меньшим разнообразием диаминокислот.
Наружная мембрана представляет собой толстый слой клеточной стенки и состоит из липополисахаридов, фосфолипидов и белков. Принципиальная структура этого слоя очень похожа на структуру плазматической мембраны. Непосредственно над муреиновым слоем расположен слой фосфолипидов, молекулы которых ориентированы своими гидрофильными головками к муреину, а гидрофобными «хвостами» — к слою липополисахаридов. Липополисахариды служат основными компонентами наружной мембраны и имеют необычное строение. Их молекулы состоят из трех частей: липида А, ядра и О-антигенов.
Липид А — «скелет» сложной молекулы — представляет собой дисахаридные звенья, образованные двумя остатками N-ацетилглюкозамина, связанными b-(1®4)-гликозидными связями, которые многократно повторяются и соединяются между собой с помощью фосфодиэфирных мостиков. К остаткам N-ацетилглюкозамина присоединяются остатки жирных кислот, которые совместно с фосфолипидами формируют липидный бислой. В двойной липидный бислой, образованный липополисахаридами и фосфолипидами, вкраплены молекулы белков. Они пронизывают бислой и представляют собой трансмембранные транспортные белки, ответственные за перенос веществ через наружную мембрану. Снаружи к липиду А присоединяются полисахаридные молекулы, которые можно разделить по длине на 2 части — ядро (включает 8— 10 остатков моносахаридов и почти у всех грамотрицательных бактерий имеет одинаковую структуру) и О-антигены. Последние представляют собой вариабельную часть наружных полисахаридов и чаще всего образованы тетрасахаридными последовательностями, которые могут повторяться в одной молекуле до 50 раз (рис. 5.7).
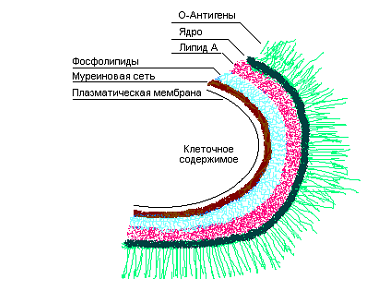
В отличие от ацильной части липида А, олигосахариды ядра и О-боковые цепи высоко гидрофильны. Ряд сахаров в липополисахариде фосфорилирован, и молекула в целом имеет отрицательный заряд. Для поддержания стабильности липополисахаридного слоя в нем присутствуют в большом количестве ионы Са2+. О-боковые цепи находятся на самой поверхности клетки и образуют так называемый «молекулярный ворс». Они служат рецепторами для адсорбции многих бактериофагов, а также являются главными антигенными детерминантами клеточной стенки грамотрицательных бактерий. Их называют, кроме того, эндотоксинами, поскольку вирулентность бактериальных клеток для животных определяется наличием и типом О-боковых цепей. В экспериментах по введению лабораторным животным выделенных клеточных стенок бактерий определенного вида происходит копирование симптомов соответствующего заболевания, и у животных вырабатывается иммунитет к бактериям — обладателям использованных клеточных стенок. Наружная мембрана соединяется с муреиновой сетью с помощью липопротеинов, которые через диаминопимелиновую кислоту присоединяются к муреину. При этом липофильные (гидрофобные) части липопротеинов погружены в липидный бислой и выполняют роль «якоря».
Ряд химических факторов может оказывать воздействие на клеточные стенки прокариот, в частности такие широко распространенные агенты, как фермент лизоцим и антибиотик пенициллин. Механизмы этих явлений будут рассмотрены далее в соответствующих разделах. Общим для действия названных факторов и им подобных является то, что в гипертонических средах оно приводит к образованию протопластов (полностью лишенных клеточной стенки форм) или сферопластов (форм, частично лишенных клеточной стенки). Эти образования характеризуются осмотической хрупкостью (лизируются в гипотонических средах) и способны в подходящих условиях регенерировать клеточную стенку. Протопласты и сферопласты широко используются в генной инженерии в экспериментах по их слиянию и трансформации с получением гибридных бактерий.
Дата добавления: 2016-05-30; просмотров: 4357;
Поиск по сайту
Узнать еще
- a-спираль b-складчатая структура
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I.2. Антигены системы АВ0. Генетика. Структура
- II.1.2. Гены. Структура антигенов
- MS Word: проверка орфографии, поиск и замена, режимы просмотра документа (обычный, разметки страницы, структура).
- VIII. Формирование и структура характера
- ІІ.5.3. Комбинаторика крестовых клеточных структур
- ІІ.5.4. Определение фрактальной размерности крестовых клеточных структур

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
