Основные принципы оптимального поведения
Поведение, которое кажется явно адаптивным, или хорошо спланированным, может быть либо результатом использования животным простейших эмпирических правил поведения, либо представлять собой когнитивное, или намеренное, поведение (см. разд. 26.7). Например, ребенок может пересечь улицу в соответствии со строгими правилами уличного движения. Если ребенка хорошо обучили, то его поведение при переходе дороги будет автоматизированным. Взрослый же человек, который не обучался этим правилам, например иностранец, будет обдумывать, как перейти улицу, оценивать скорость и характер движения приближающегося транспорта и т. д. Внешняя картина поведения ребенка и взрослого при переходе через улицу может быть практически неотличимой, но в одном случае это поведение осуществляется на основе простейших эмпирических правил, а в другом - на основе познавания.
Можно обеспечить оптимальность поведения посредством простого набора правил. Пример такого рода мы находим в работе Грина (Green, 1983), проанализировавшего правила остановки, которые должны обеспечивать оптимальность пищедобывательного поведения. В своей работе Грин предположил, что животные-жертвы распределены по различным участкам земли, которые различаются по качеству, и на лучших из них хищники гораздо быстрее ловят свою добычу. В разных условиях среды распределение участков по качеству будет различным. Предполагается, что хищник в состоянии различать типы кормных участков, только оценивая свой успех на каждом из них. Хищник не возвращается на тот участок, где он уже побывал, и систематически обследует каждый участок до тех пор, пока не решит оставить его и перебраться на другой.
Оптимальную стратегию пищедобывания можно охарактеризовать правилом остановки, которое определяет, когда именно хищнику следует оставить данный участок. В любое время хищник может решить, уйти или остаться на данном участке, чтобы продолжать поиски жертвы. Грин показывает, что наилучшее правило остановки - это то, в основу которого положено количество добычи как функция времени, потраченного на обследование данного участка. Альтернативные правила остановки включают в себя: наивную стратегию, при которой хищник полагается на знание средней вероятности обнаружения жертвы на каждом участке; всеведущую стратегию, при которой хищник может оценить качество каждого участка, не обследуя его, и таким путем может избежать бедных добычей участков, и, наконец, стратегию учета мгновенной скорости пищедобывания. При этой стратегии хищник покидает участок охоты, когда эта скорость падает ниже критического уровня. Наилучшая стратегия, по Грину, подразумевает оценку качества участка по мере его обследования. Эта стратегия продуктивнее, чем наивная стратегия и стратегия учета мгновенной скорости пищедобыва-
ния. Она также более продуктивна, чем всеведущая стратегия, потому что предъявляет меньше требований к способности отдельного животного производить расчеты. Стратегию Грина можно представить в виде простого правила: оставаться на участке столько времени, пока больше половины обследованных мест приносит добычу, в противном случае - уходить. Эта стратегия может быть осуществлена посредством простого механизма.
Вааге (Waage, 1979) предположил, что существует простой механизм принятия решения, которым пользуются паразитирующие осы в поисках подходящего хозяина, на которого они могли бы отложить яйца. Прилетев на подходящее место, оса начинает разыскивать нужных ей животных; ее реактивность (тенденция повернуть назад внутрь обследуемого участка при достижении его границы) при этом характеризуется каким-то начальным уровнем, устанавливающимся на основе химического стимула, выделяемого на этом участке животными-хозяевами. Такой уровень реактивности осы со временем линейно уменьшается до тех пор, пока не достигнет некоторого порогового уровня или пока оса не столкнется с искомым животным. После каждого откладывания яйца уровень реактивности осы возрастает на величину, которая зависит от времени, прошедшего после предыдущей кладки. Этот процесс продолжается до тех пор, пока реактивность в конце концов не снизится до пороговой величины, и тогда оса улетает с этого участка.
Модели Грина (Green, 1980; 1983) и Вааге (Waage, 1979) дают сходные результаты. Однако при этом важно помнить, что у Грина это функциональная модель, которая точно определяет, что именно животному следует делать, чтобы достигнуть наилучшего результата. Модель Вааге механистическая, и она построена на основе представлений о непосредственных причинах возникновения поведения.
Один из методов, с помощью которого можно определить, следует ли животное в выборе своего решения тем или иным фиксированным правилам, состоит в том, чтобы каким-то образом избирательно вмешиваться в его поведение. Например, при исследовании поведения роющих ос (Ammophila campestris) Берендс (Baerends, 1941) обнаружил, что перед тем, как отложить яйцо, самка роет норку, убивает или парализует гусеницу бабочки, несет ее к норке, откладывает на гусеницу яйцо и прячет ее в норке. Эту процедуру самка осы повторяет затем при откладывании второго и каждого последующего яйца. Тем временем созревает первое яйцо, и личинка начинает пожирать гусеницу. Теперь оса возвращается к первой норке и добавляет в нее новых гусениц. После этого в зависимости от обстоятельств она может приступить к изготовлению новой норки или будет снабжать гусеницами вторую норку. Таким образом самка осы может обслуживать до пяти гнезд одновременно (рис. 25.16).
Берендс обнаружил, что осы каждое утро проверяют все норки, прежде чем отправиться в свои «охотничьи угодья». Забирая гусениц из норки, Берендс мог заставить осу приносить больше пищи, чем обычно; добавляя гусениц, он мог заставить ее приносить меньше пищи. Однако он мог таким образом управлять поведением осы только в том случае, если производил изменения в гнезде перед первым ежедневным посещением норки осой. Если же такие изменения совершались в течение дня после этого момента, они не вызывали никакого эффекта. По-видимому, самка осы руководствуется какими-то простыми правилами. Существует стандартный порядок действий, необходимых для откладывания яйца, который предусматривает рытье норки и заготовку гусеницы. Кроме того, имеется стандартный распорядок проверки ранним утром всех норок, в ходе которого обычно устанавливается, в какое гнездо в течение дня необходимо принести пищу. И наконец, существует стандартный порядок прекращения этой деятельности, в соответствии с которым оса закрывает гнездовую норку, когда в ней окажется достаточное число гусениц. Хотя она и способна при посещении гнезда оценить количество запасенной в нем пищи, она не всегда использует эту способность. Более того, каждая из стандартных последовательностей действий, будучи начата, продолжается до полного завершения. Так, например, оса будет приносить и прино-
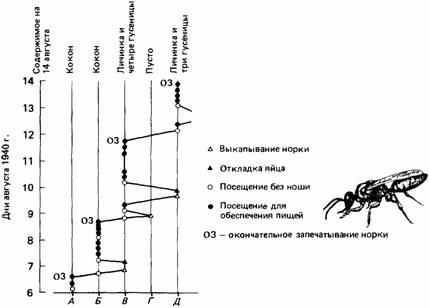
Рис. 25.16.Диаграмма гнездового поведения роющей осы (Ammophila). (По Baerends, 1941.)
сить в гнездо гусениц, если их систематически каждый раз удалять из гнезда, как только оса их принесет. Этот пример показывает, что сложное поведение может быть запрограммировано в виде набора жестких правил. Оса ведет себя наподобие автомата, хотя у нее и есть некоторые стандартные программы поведения, позволяющие ей выходить из затруднительного положения, например удалять из норки какие-то препятствия.
Как мы уже видели ранее, прерывание поведения животного при определенных обстоятельствах маскирует поведение, которое должно было бы проявиться, если бы не было этого перерыва. Такая ситуация разделения времени предполагает, что животное следует определенным правилам, которые определяют организацию и приоритетность поведенческих актов в общей картине поведения. Рассмотрим конкретный пример. Когда голодная горлица (Streptopelia) ест, либо выбирая зерна из кучи, либо получая пищу в камере Скиннера, в ее поведении можно наблюдать типичные паузы длительностью в несколько минут (см. рис. 25.11). Что будет делать горлица во время этих пауз, зависит от обстоятельств. Если птица имеет доступ к воде, она будет пить. В противном случае она может чистить перья или просто неподвижно стоять. В экспериментальных условиях было показано, что на время возникновения этих пауз никакого влияния не оказывает манипулирование мотивационными факторами вторичного приоритета, например изменением уровня жажды. В одном из экспериментов голодным горлицам к каждому крылу прикрепили по канцелярской скрепке. Во время еды горлицы не обращали на скрепки никакого внимания, тогда как во время пауз они пытались от них отделаться. Однако наличие канцелярских скрепок не оказывало влияния на характер пищевого поведения и не меняло временного распределения пауз (McFarland, 1970b). Создается впечатление, что в пищевом поведении горлицы как бы запрограммированы на строго определенное время паузы и что правила, которые регулируют пищевое поведение горлиц, не находятся под каким бы то ни было влиянием со стороны других мотивационных факторов, например жажды или стремления чистить перья, если только эти тенденции не станут сильнее, чем пищевая. Это типичный случай явления разделения времени.
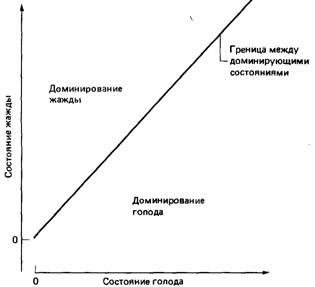
Рис. 25.17. Граница между состояниями животного, в которых доминируют голод и жажда.
Если каким-то путем прервать пищевое поведение голодной горлицы, то обычно после перерыва она продолжит это поведение. Но если прервать процесс питья воды, то, как правило, он будет маскироваться, если перерыв будет достаточно большим (McFarland, Lloyd, 1973). В экспериментальной ситуации с инструментальным поведением, где горлицы для получения пищи и воды должны клевать светящиеся клавиши, прерывания текущей деятельности можно добиться простым выключением подсветки клавиши. Птицы быстро научатся прекращать клевание, когда эти клавиши не светятся. В условиях свободного пищевого и питьевого поведения прерывания поведения можно достигнуть, если экспериментальную комнату примерно на минуту погрузить в темноту. При сравнении оказалось, что эти два типа прерывания деятельности горлиц оказывают одинаковый эффект (Larkin, McFarland, 1978).
Разделение времени при пищевом и питьевом поведении горлиц послужило объектом многочисленных экспериментов, целью которых было обнаружить правила, на основе которых птица принимает решение, есть ей или пить. Результаты показывают, что, во-первых, в экспериментах может доминировать либо питьевая, либо пищевая деятельность (McFarland, Lloyd, 1973; McFarland, 1974). Во-вторых, линия, отражающая границу (рис. 25.17) между доминированием голода и доминированием жажды, не меняет своего положения ни при повторных опытах, ни при различных начальных уровнях голода и жажды животного, ни при изменении результатов пищевого и питьевого поведения (Sibly, McCleery, 1976). Однако если в ходе эксперимента изменять мотивационное состояние птицы, то может показаться, что произойдет поворот графика, отражающего границу между доминирующими состояниями (рис. 25.18). Теоретический анализ этой ситуации показывает, что никакого реального изменения в положении границы доминирующих состояний не происходит. Это кажущееся изменение обусловлено используемой экспериментатором системой координат, потому что мотивационное состояние животного обычно изображается в двух измерениях, тогда как здесь необходимо учитывать и другие измерения (McFarland, Sibly, 1975). Величина этого кажущегося поворота границы доминирующих состояний оказалась полезным показателем для измерения силы мотивационных факторов, таких, как величины привлекательности пищевого и питьевого подкреплений (Sibly, 1975), эффективность внешних стимулов, которые сиг-
нализируют о доступности пищи и воды (McFarland, Sibly, 1975; Beardsley, 1983), и затраты (оцениваемые самой птицей) на смену пищевого поведения на питьевое и наоборот (Larkin, McFarland, 1978). В целом создается впечатление, что и внутренние, и внешние факторы оказывают какое-то влияние на. тенденции пищевого и питьевого поведения и что эти тенденции конкурируют за доминирование (McFarland, 1974). Заняв доминирующее положение, победившая система периодически предоставляет время для осуществления другой (субдоминантной) деятельности. Почему поведение организовано именно таким образом, остается загадкой.
Вполне возможно, что в пищевом поведении горлиц паузы представляют собой часть стратегии поведения, направленной на обнаружение хищников. Находясь в стае, отдельные птицы имеют возможность тратить больше времени на добывание пищи и меньше - на то, чтобы следить, не появится ли хищник (Barnard, 1980; Bertram, 1980; Elgar, Catterall, 1981). Лендрем (Lendrem, 1983) обнаружил, что горлицы, питаясь в одиночку, тратят около 25% времени из двухминутного периода пищевого поведения на то, что оглядываются по сторонам, а когда рядом есть другие птицы - около 20%. Однако эта разница была гораздо более выраженной, если горлицы незадолго до этого видели поблизости хищника (хорька). В этой ситуации одинокие горлицы тратят около половины времени на то, чтобы осматриваться вокруг, тогда как в присутствии двух других птиц у них уходит на это только 25% времени. Время, затрачиваемое не на добывание пищи, уменьшалось еще больше, когда число птиц в стае возрастало. По мере увеличения размера стаи горлицы получали пищу быстрее, в то же время общая скорость добывания пищи снижалась, если они недавно видели хищника. Как показывает детальный анализ пищевого поведения горлиц, в ситуациях риска у них снижается скорость добывания пищи, тогда как паузы между приемами пищи увеличиваются. Таким образом, они питаются медленнее, когда находятся одни в незнакомом окружении и вскоре после того, как видели хищника. Особенно увеличивается при этом период времени после каждого клевка, когда горлица стоит с поднятой головой; возможно, что это повышает способность птицы заметить хищников.
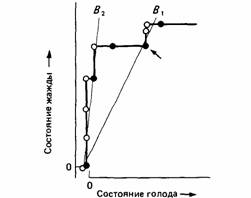
| Рис. 25.18. Кажущийся поворот границы между доминирующими состояниями. В результате экспериментальных манипуляций (обозначено стрелкой) создается впечатление, что граница сдвигается из положения В1в положение В2 . Новое положение границы определяли в эксперименте, в котором все субдоминантное поведение максировали прерыванием. Черными кружками обозначены состояния, в которых доминирует голод, а белыми - в которых доминирует жажда. (По McFarland, 1974.) |
Скорость добывания пищи снижается также и в том случае, когда горлицам приходится отличать пригодную пищу от непригодной. Добавляя хлорид лития к пище горлиц в сочетании с определенными условиями кормления, можно научить этих птиц, как и многих других животных, избегать пшеничных зерен, окрашенных в определенный цвет (Lendrem, McFarland, 1985). Наученные птицы ведут себя таким образом, как если бы зерна этого цвета были ядовитыми. Например, одни птицы избегают желтых зерен, а другие - красных. Когда таким горлицам дают смесь красных и желтых зерен, им приходится различать эти два типа зерен, чтобы избежать зерен того цвета, к которому у них образовалось отвращение. Птицы, которые добывают пищу из смеси «ядовитых» и безвредных зерен, питаются медленнее, чем птицы, которым дается смесь из «неядовитых» зерен различного цвета (Lendrem, McFarland, 1985).
Если горлица питается медленнее обычного, поскольку ей приходится отличать безвредные зерна от ядовитых, то можно думать, что у нее ослаблена способность обнаруживать хищников, поскольку она обращает больше внимания на пищу. На самом же деле скорость реагирования горлиц на пролетающую над их головой модель ястреба повышается, если горлицам дают смесь ядовитых и безвредных зерен (Lendrem, McFarland, 1985). Птицы, которым предварительно показали хищника (и которые поэтому клюют зерна с пониженной скоростью), быстрее реагируют на модель ястреба, чем птицы, которым приходится отличать безвредную пищу от ядовитой. Таким образом, создается впечатление, что более медленное питание, какой бы причиной оно не вызывалось, увеличивает способность птицы обнаруживать хищников. Эти данные согласуются с представлением о том, что высокая скорость добывания пищи (или другого поведения) связана с большими затратами.
Что же произойдет, если мы еще усложним задачу по различению зерен, положив их на таком фоне, где они будут плохо различимы? Как и следовало ожидать, произойдет дальнейшее снижение скорости пищевого поведения (рис. 25.19). Отчасти это может быть связано с тем, что птицы должны обращать больше внимания на добывание пищи, но это может быть и проявлением активной тактики, направленной на то, чтобы не терять бдительности. Горлицы, которые выбирают безвредные зерна из смеси с ядовитыми на таком фоне, где они плохо различимы, быстрее замечают модель ястреба, чем птицы, выбирающие хорошо различимые зерна (рис. 25.20) (Lendrem, McFarland, 1985). Однако птицы, которые выбирают подходящие зерна в условиях низкой их различимости, совершают больше ошибок (едят больше ядовитых зерен) и реже делают паузы в пищевом поведении, чем птицы, питающиеся хорошо различимыми зернами. Таким образом, совершенно очевидно, что существует определенный баланс между требованиями бдительности и требованиями пищедобывания.
В заключение следует сказать, что, по всей вероятности, горлицы, которые по-
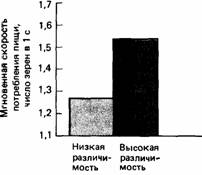
Рис. 25.19.Скорость поглощения пищи горлицами, которым была предложена смесь «ядовитых» и безвредных зерен, в условиях, когда зерна этих двух типов трудно (низкая различимость) или легко (высокая различимость) отличить друг от друга. (Lendrem, McFarland, 1985.)
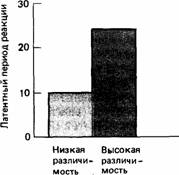
Рис. 25.20.Латентные периоды реакции на модель ястреба у горлиц, которые питаются в условиях низкой и высокой различимости пищи (рис. 25.19). Обратите внимание на то, что горлицы в условиях низкой различимости зерен хотя и питаются медленнее, зато быстрее реагируют на потенциального хищника. Эти результаты позволяют предположить, что пониженная скорость поглощения пищи при поедании плохо различимого зерна обусловлена не тем, что для различения зерен птица должна сосредоточить на этом все внимание, а скорее тем. что эта ситуация более опасна (вследствие повышенной вероятности проглотить «ядовитое» зерно), и поэтому горлицы обращают больше внимания на окружающую обстановку, в целом. (По Lendrem, McFarland, 1985.)
едают пищу быстро, имеют меньше шансов заметить хищников. Когда горлицы настороженны, т. е. когда они находятся в незнакомой обстановке, или в одиночестве, или в ситуации, где они недавно видели хищника, горлицы поедают пищу медленнее. Однако у горлиц существует целый ряд различных способов, с помощью которых они могут снизить общую скорость поедания пищи. Например, они имеют возможность чаще делать паузы, удлинять их или уменьшать скорость собственно поедания пищи. Эти способы могут увеличить шансы заметить хищника. Есть некоторые указания на то, что эти различные методы компенсируют друг друга (Lendrem, McFarland, 1985). Вполне возможно,
что горлицы полагаются на то, что смогут обнаружить необычное движение во время подъема головы после каждого клевка, а паузы делают, чтобы время от времени оглядеться вокруг. Не исключено, что, делая паузы в клевании, птица может потратить некоторое время на чистку перьев или питье, - это пример феномена, называемого разделением времени. В настоящее время мы не обладаем достаточными знаниями о зрении птиц, чтобы подтвердить эти гипотезы. Мы также не знаем, используют ли птицы какой-то сложный набор правил принятия решений или же их поведение регулируется посредством когнитивных процессов.
ДЛЯ ЗАПОМИНАНИЯ
1. Животные могут принимать решения на основе простых эмпирических правил, которые помогают им приспособиться к конкретным особенностям окружающей среды.
2. Если при манипулировании второй по приоритету активностью изменяется распределение во времени переключений животного с одной деятельности на другую, то можно сделать вывод, что эти переключения обусловлены конкуренцией мотиваций. Если же это распределение не меняется, то такие переключения вызваны растормаживанием.
3. В том случае, когда момент начала и продолжительность проявления какой-то деятельности регулируются другой деятельностью, можно говорить, что поведение организовано в режиме разделения времени.
4. Принятие оптимального решения животным реализуется в последовательности поведенческих актов, которая максимизирует некоторый показатель приспособленности организма при существующих условиях. Любое нарушение взаимного соответствия между животным и его средой обитания будет приводить к тому, что такая максимальная приспособленность будет редко достигаться. Однако животные могут использовать такие правила принятия решения, что их поведение будет близким к оптимальному.
Рекомендуемая литература
McFarland D.J. (1977). Decision-making in animals, Natura (London), 269, 15-21.
Krebs J. R., McCleery R. //., 1984. Optimisation in behavioural ecology. In: Krebs J. R., Davies N. B. (eds), Behavioural Ecology, 2nd edn, Oxford, Blackwell Scientific Publications.
Дата добавления: 2016-06-18; просмотров: 2620;
Поиск по сайту
Узнать еще
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cущность организации и ее основные признаки
- F68 Другие расстройства зрелой личности и поведения у взрослых
- I. Назначение унифицированных газозарядных станций и основные тактико-технические требования, предъявляемые к ним.
- I. Общие принципы структурно-функциональной организации клетки и её компоненты. Плазмолемма, её структура и функции.
- I. ОСНОВНЫЕ ПОЛОЖЕНИЯ
- I. Политический режим: понятие, сущность и основные типы.
- I. Этические принципы психолога

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
