Конкуренция мотиваций
Исходя из простой теории конкуренции мотиваций, можно было бы ожидать, что находящееся в состоянии голода и жажды животное будет вести себя таким образом, как показано на рис. 25.4, т.е. пить оно станет тогда, когда жажда у него будет сильнее голода, а есть - когда голод будет сильнее жажды. Мы должны были бы ожидать, что животное будет вести себя рационально в том отношении, что его предпочтения будут транзитивными. Вместе с тем при анализе рис. 25.4 мы видим, что весьма маловероятно, чтобы животное было организовано таким образом, т.е. чтобы оно проявляло подобное поведение. Линия ОХ на этом рисунке обозначает совокупность таких состояний животного, когда голод и жажда его одинаковы по силе. Вблизи этой линии график состояния животного должен был бы все время колебаться, так как животное должно было бы все время переключаться с одного поведения на другое. Если у животного голод чуть-чуть сильнее жажды, то проглоченный кусочек пищи уменьшит голод, и теперь уже жажда окажется сильнее. Один глоток воды восстановит прежнюю ситуацию. Такие колебания не могут быть эффективной формой поведения, поэтому был выдвинут целый ряд других предположений относительно механизмов организации поведения. Когда инженеры сталкиваются с подобной проблемой при конструировании каких-либо машин, то для предотвращения колебаний они иногда используют гистерезис. Гистерезис - это некий механизм, обеспечивающий задержку между стимулом и реакцией на него. Высказывалось предположение (например, Toates, Oatley, 1970; Toates, 1980), что гистерезис - это компонент механизма, регулирующего пищевое и питьевое поведение животных. Наличие гистерезиса приводит к тому, что животное ощущает уменьшение чувства голода не сразу, как только начнет есть, а лишь значительное время спустя. В рассматриваемом примере действие гистерезиса будет проявляться в том, что траектория переключений между проявлениями голода и жажды будет заходить за разделительную линию, соответствующую равенству мотиваций голода и жажды.
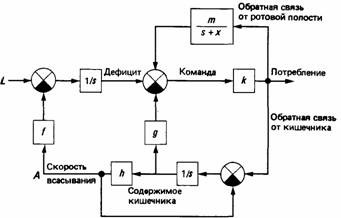
Другой возможный механизм, который оказывает почти такой же эффект, - это положительная обратная связь. Имеются некоторые данные, свидетельствующие о том, что стремление есть (Wiepkema, 1971) или пить (McFarland, McFarland, 1968) временно усиливается в результате прохождения пищи или воды через ротовую полость. Эта положительная обратная связь от приема пищи и воды существует параллельно с обычной отрицательной обратной связью, которая ведет к насыщению (рис. 25.5). Так, например, если горлице заливать воду через трубочку прямо в зоб, то наступит утоление жажды точно так же, как и при обычном питье воды. Однако в ситуации с инструментальным поведением поступающая через трубочку вода не даст подкрепляющего эффекта. Прохождение воды через ротовую полость действует как положительное подкрепление, но никак не
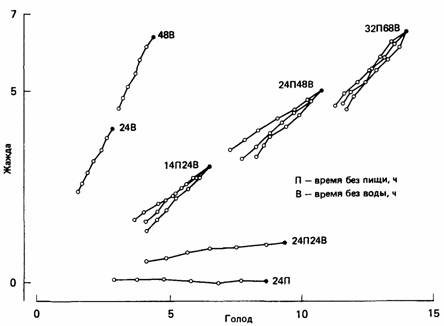
Рис. 25.3. Горлицам, которые находились в различных исходных состояниях (черные кружки) голода и жажды, через каждые 30 мин предлагали осуществить выбор между пищей и водой. Суммарное потребление горлицами пищи и воды в течение получасового эксперимента показано белыми кружками (один кружок через каждые 5 мин). Обратите внимание на то, что при осуществлении выбора горлицы отнюдь не стремятся уравнять голод и жажду, как можно было бы ожидать, если бы их поведение определялось простой конкуренцией мотиваций (рис. 25.4). (McFarland, 1971.)
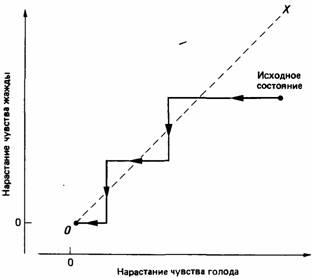
Рис. 25.4.Схематическое изображение простой конкуренции мотиваций. Точки пространства, расположенные справа от штриховой линии, обозначают состояния животного, когда голод сильнее, чем жажда, а слева - состояния, когда жажда сильнее, чем голод. В соответствии с теорией прортой конкуренции мотиваций животное сначала удовлетворяет более сильную мотивацию до тех пор, пока она не станет равна второй имеющейся у него мотивации. Затем животное поочередно реализует то одну, то другую мотивацию (величина ступенек на графике сильно преувеличена) до тех пор, пока обе мотивации не уменьшатся до нуля.
Рис. 25.5. Блок-схема механизма управления пищевым поведением у горлиц, в которой сигналы, исходящие от ротовой полости, обеспечивают положительную обратную связь, а сигналы от кишечника - отрицательную обратную связь (кружками с крестами обозначены суммирующие узлы, причем черный квадрант в кружке означает изменение знака сигнала). L - скорость потери воды; А - скорость всасывания воды в кровь. Другими символами обозначены параметры составных частей управляющего механизма. (McFarland, McFarland, 1968.)
способствует утолению жажды (McFarland, 1969b). Эффект положительной обратной связи будет проявляться в том, что график состояния животного будет отклоняться в стороны от линии, соответствующей равенству мотиваций голода и жажды. Совершенно очевидно, однако, что истинное положение дел гораздо сложнее. Я обнаружил, что если горлицы получали еду и питье в таких условиях, где для смены пищевого поведения на питьевое и наоборот им приходилось преодолевать определенные препятствия, то в случае больших препятствий они реже производили такую смену активности, чем в случае малых (рис. 25.6). Отсюда следует, что горлицы, по-видимому, принимают решение о том, когда именно переключиться с одной активности на другую, с учетом трудностей или затрат, связанных с таким переключением (см. гл. 24).
Теперь мы видим, что очень маловероятно, чтобы в основе механизма, определяющего принятие животным решения о смене одного поведения на другое, лежала простая конкуренция соответствующих мотиваций. Вместе с тем не исключено, что какая-то более изощренная версия механизма конкуренции мотиваций могла бы дать необходимый набор правил принятия решения. Как мы уже знаем из гл. 15, мотивационное состояние животного может быть результатом действия самых различных факторов, таких, как оценка животным внешних стимулов (силы сигнальных раздражителей), его исходное внутреннее состояние и, возможно, вторичные мотивационные факторы, как, например, скорость изменения состояния. Совместное действие всех этих факторов создает общее мотивационное состояние, которое можно представить в виде движущейся точки (или траектории) в пространстве мотивационных состояний. Различные состояния могут порождать одну и ту же поведенческую тенденцию, и соответствующие точки можно соединить, получив при этом мотивационную изоклину. Исходя из этого, мы можем себе представить, что существует простое правило конкуренции для выбора одной из тенденций к поведению, но при этом любая тенденция представляет собой результат взаимодействия разнообразных факторов, каждый из которых тщательно взвешен с точки зрения того, насколько он отражает самые главные интересы животного. Например, Сибли (Sibly, 1975) полагает, что, когда горлицам приходится выбирать между пищевым и питьевым поведением в их инструментальной деятельности, они руководствуются следующим правилом принятия решения: «Ешь, если произведение потребности в пище на пищевую привлекательность больше, чем произведение потребности в воде на питьевую привлекательность. Пей, если произведение потребности в пище на пищевую привлекательность
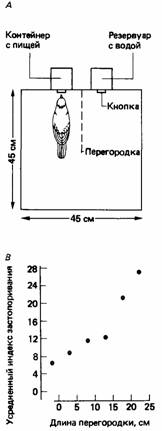
| 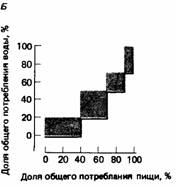 Рис. 25.6. А. Вид сверху на камеру Скиннера, в которой горлицы могли получить пищу и воду, поклевав соответствующую кнопку. Длину прозрачной перегородки между кнопками можно было изменять. Б. Жирной линией показан ход пищевого и питьевого поведения горлицы вплоть до полного удовлетворения этих мотиваций (100%). Обратите внимание на то, что птица четыре раза переключалась с одного поведения на другое и обратно. Подсчитав суммарную площадь заштрихованных областей на этом графике, мы получаем «индекс застопоривания», обратно пропорциональный числу переключений с одной активности на другую. В. Зависимость усредненной величины индекса застопоривания от длины перегородки. (McFarland, 1971.)
Рис. 25.6. А. Вид сверху на камеру Скиннера, в которой горлицы могли получить пищу и воду, поклевав соответствующую кнопку. Длину прозрачной перегородки между кнопками можно было изменять. Б. Жирной линией показан ход пищевого и питьевого поведения горлицы вплоть до полного удовлетворения этих мотиваций (100%). Обратите внимание на то, что птица четыре раза переключалась с одного поведения на другое и обратно. Подсчитав суммарную площадь заштрихованных областей на этом графике, мы получаем «индекс застопоривания», обратно пропорциональный числу переключений с одной активности на другую. В. Зависимость усредненной величины индекса застопоривания от длины перегородки. (McFarland, 1971.)
|
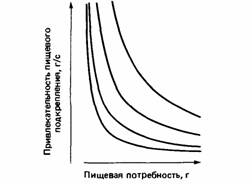
меньше, чем произведение потребности в воде на питьевую привлекательность». Сибли определяет привлекательность в понятиях скорости, с которой могли бы быть получены пища и вода. Такие правила принятия решения соответствуют конкуренции между пищевой и питьевой тенденциями, каждая из которых основана на гиперболических мотивационных «изоклинах» (рис. 25.7).
Можно допустить, что основное правило принятия решений, которым пользуются животные, базируется на некоторой форме конкурентных отношений. Когда же мы встречаемся с нарушениями этого правила, то их можно объяснить, предполагая, что животному приходится соответствующим образом выверять и взвешивать различные факторы, которые объединяются и порождают тенденцию для выполнения определенного типа поведения. Это спорная тема, и аргументы здесь основаны на очень сложном жонглировании различными переменными и параметрами (например, Houston, 1982; McFarland, 1983). Тем не менее студенты должны помнить несколько общих принципов. Во-первых, любая теория, которая утверждает, что самая сильная тенденция всегда одерживает верх при конкуренции и, следовательно, определяет поведение животного, таит в себе опасность тавтологии. Она подразумевает, что наблюдаемое поведение всегда отражает самую сильную тенденцию. Если такая теория фактически приравнивает поведение к тенденции, то она бесполезна для объяснения процесса принятия решения у животных. Во-вторых, встает вопрос о том, будет ли иметь какой-либо биологический смысл механизм принятия решения, если он основан только на конкуренции мотиваций. Не будет ли животное пре-
|
| Рис. 25.7. Изоклины гиперболической формы, соединяющие точки, где пищевая тенденция имеет одинаковую величину. Графики построены исходя из гипотезы о том, что пищевая тенденция есть произведение потребности в пище на ее привлекательность. |
небрегать некоторыми низкоприоритетными видами поведения, например чисткой шерсти? Будет ли способно животное корректировать желаемую последовательность поведенческих актов под давлением обстоятельств, что кажется вполне разумным с эволюционной точки зрения? В-третьих, существует ли какая-либо альтернатива конкурентной теории принятия решения? Это трудный вопрос. С одной стороны, можно утверждать, что конкуренция мотиваций — это необходимый аспект рационального выбора альтернатив, которые не могут одновременно проявиться (например, Ludlow, 1980). С другой стороны, можно представить себе и нерациональное принятие решения, подобное тому, что мы уже видели ранее при обсуждении циклического шаблонного поведения пескожила, которое тем не менее адекватно обстоятельствам, в которых живет это животное.
Дата добавления: 2016-06-18; просмотров: 2859;
Поиск по сайту
Узнать еще
- Конкуренция «газ-газ»
- Конкуренция больших и маленьких
- Конкуренция и результативность
- Конкуренция с ядом за связывание с ферментом
- Конкуренция, её формы и методы. Монополия и конкуренция.
- Конкуренция: сущность и виды. Формы конкурентной борьбы.
- Конкуренция: сущность, виды и формы конкурентной борьбы.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
