Баланс вещества и энергии
Поскольку основной задачей физики ландшафта является изучение перемещений и превращений вещества и энергии, среди методов научного обобщения материалов особое значение приобретает метод балансов (Арманд, 1947а).
Этот метод позволяет оценивать количество различных форм вещества и энергии, поступающих в ландшафт и выходящих из него, прослеживать динамику суточных и годовых циклов, анализировать распределение потоков вещества и энергии по разным каналам в зависимости от типа ландшафта, вскрывать тенденцию вековых изменений.
Наиболее разработаны, хотя еще далеко несовершенны, методы составления трех взаимосвязанных балансов: радиационного, теплового и водного. Однако ими не исчерпываются возможности балансового метода в применении к абиотическим компонентам ландшафта. Энергетические потоки, проходящие через балансируемый объем, не ограничиваются солнечной радиацией и длинноволновым излучением.
Возможно составление баланса кинетической энергии ветра, энергии водяного пара и облаков, как потенциальной, связанной с их высотным положением, так и тепловой, связанной с термическим и фазовым состоянием водных запасов атмосферы. Количественно, в энергетических терминах, еще не изучено взаимодействие ветра с подстилающей поверхностью: гашение энергии ветра, изменение испарения и транспирации, дефляция, перевевание снега.
Огромная суммарная энергия падающего дождя не может не оказывать влияния на растительность и верхний горизонт почвы. В массе осадков остается запас потенциальной энергии, пропорциональный альтитудам мест выпадения. Этот запас преобразуется в энергию потоков, не только производящих эрозию и транспортирующих наносы, но и освобождающих, подобно реле, огромную потенциальную энергию горных пород и почвы, покоящихся над базисом денудации.
Исследование балансов энергии ветра и атмосферной влаги не покажется слишком отвлеченным, если мы учтем приближение эпохи управления климатом - задачи именно энергетической, требующей точного знания мощности как катализаторов, так и подлежащих их воздействию воздушных масс.
Наименее разработан баланс биомассы. Здесь сами измеряемые параметры очень сложны и множественны (Арманд, 1949а). Если говорить о фитомассе, то ее годовой приход устанавливается путем периодического взвешивания единовременного запаса. Но годовой так называемый брутто- приход не равен разности между максимумом и минимумом годового хода, но всегда больше его, так как, кроме разового осеннего увядания, непрерывно происходят расход живой растительной ткани на дыхание, частичное отмирание органов (опад), поедание растений животными и преждевременное умирание растений по какой-либо причине (отпад).
И если считать годовым приходом не указанную разность (нетто-приход), а всю сумму непрерывно синтезируемой фитомассы, или статьи расхода должны учитываться.
Годовой баланс многолетних и однолетних фитоценозов принципиально различен. В подрастающем лесу годовая балансовая разность всегда положительна, при достижении спелости и нормальном возобновлении она становится нейтральной, при возникновении неблагоприятных условий лес деградирует и разность становится отрицательной.
Но во всех случаях в лесу годовой прирост фитомассы намного меньше единовременной массы, в составе которой подавляющую роль играет переходящий запас древесины. В однолетних ценозах, например полевых культурах, годовой минимум равен нулю. Балансовая разность также всегда равна нулю, но в данном случае по этому признаку уже нельзя судить о стабильности ландшафта; тенденция развития может быть вскрыта лишь сравнением годовых максимумов (урожаев).
Таким образом, для сообщества однолетних растений значимый годовой прирост положительной и отрицательной частей баланса всегда связан с изменением условий среды (свет, тепло, влага, углекислота, питательные вещества почвы, трофические связи), для сообщества многолетних — как с изменением условий, так и с возрастом.
Динамика естественного травостоя соответствует динамике однолетних культур лишь в том случае, если мы условно абстрагируемся от массы перезимовывающих подземных органов растений ввиду чрезвычайной трудности ее учета. В действительности фитомасса травостоя изменяется по смешанному типу, причем подземные части многолетников и семена однолетников играют роль переходящего запаса.
Единовременный запас фитомассы обычно измеряется в единицах сухого веса. Однако для годового прихода измерение в этих единицах приобретает несколько фиктивный характер, так как масса прихода, постоянно несущая потери, никогда одновременно в полном объеме не существует.
В этом случае более правильно пересчитывать приход в весовые единицы углерода, связанного за балансовый период в ходе фотосинтеза. Но и этот способ несовершенен. Во-первых, при отмирании растений не происходит немедленного освобождения углерода хотя бы до формы окисла, а осуществляется лишь постепенный переход его в более простые органические соединения; в балансе появляется промежуточная категория мертвой органики - опад, в свою очередь проходящий на пути к минерализации ряд стадий.
Во-вторых, в процессе как фотосинтеза, так и последующей ассимиляции углерод переходит в более или менее высокомолекулярные соединения, так что то же количество С может создать разную биомассу в зависимости от количества О, Н, N, Са и других атомов, входящих в молекулы клетчатки, сахаров, кислот, белков, элементоорганических соединений и т.д.
Качественные различия создаваемой фитомассы могут быть учтены количественно при переходе к энергетическому балансу, т.е. при учете в калориях энергии, поглощаемой сообществом при синтезе органического вещества и освобождающейся при распаде растений.
Ценность такого метода станет очевидной, если вспомнить, что, с одной стороны, эндотермические реакции синтеза требуют тем больше энергии, чем более высокомолекулярные соединения при них возникают, с другой - именно эти соединения имеют наибольшую питательную или кормовую ценность, недаром именуемую калорийностью.
Полное потребление энергии фитоценозом не ограничивается расходом на фотосинтезе. Ход дальнейших реакций, переводящих органические вещества на более высокий энергетический уровень, десукция и транспирация также требуют расхода энергии и сильно зависят от термического режима биотопа.
Для изучения динамики ландшафта, т.е. быстроты пропускания неорганического вещества через организмы, весьма важно понятие оборачиваемости биомассы. В применении к фитомассе один из способов ее выражения - это формула, аналогичная формуле оборачиваемости капитала выражающей скорость оборота (в днях):
Т = 365 М/Ф (1), где Ф - полная продукция фитоценоза за цикл, всегда равный году; М — многолетний запас, т.е. фитомасса, содержащаяся в стволах, ветвях и корнях деревьев, корневищах, луковицах, неотмирающих корнях и семенах трав. Формула (1) дает основную продолжительность оборота, а формула (2) — число оборотов в год:
N = Ф/М (2), Нетрудно заметить, что если Ф>М, что возможно у травянистых фитоценозов, то Т получается меньше года, a N - больше единицы. Наоборот, если Ф < М, что типично для леса, то Т получается больше года и часто равно нескольким годам, a N равно правильной дроби.
Если подходить с точки зрения среды, вовлекаемой в оборот, то следует принимать Ф = П + О1 + 02 + Д, где П - нетто-продукция фитомассы, минус семена; O1 - опад и отпад в течение вегетационного периода и поедание животными однолетних частей растений; 02 — то же, многолетних; Д - потери массы на дыхание (рис. 1). Если учитывать только устойчивые продукты полураспада, например гумус, то газообразную фракцию Д принимать во внимание не нужно.
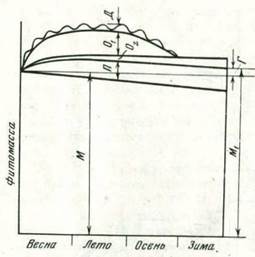
Рис. 1. Годовая динамика фитомассы в многолетнем растущем ценозе. Г - годовой прирост многолетних частей. В развивающемся ценозе M1 > М и Г > О, в достигшем спелосте ценозе М = М1 И Г = O1, в умирающем ценозе М < М1 и Г < O
Измерения динамики зоомассы во многом подобны измерениям фитомассы. Здесь годовой приход и расход также значительно больше разности между максимальным (летним) и минимальным (зимним) единовременным запасом, так как много животных погибает во все сезоны.
Переходящий остаток мал, так как наибольшей массой обладают группировки низших животных — насекомых, червей, простейших, в основном погибающих при наступлении зимы. Поскольку у наземных беспозвоночных, составляющих большую часть биомассы, жизненный цикл в подавляющей части случаев меньше года, то оборачиваемость в общем велика; она увеличивается от высших типов и классов к низшим.
Если процесс фитосинтеза и минерального питания растений всегда приводит к приросту суммарной биомассы в данном ландшафте, то при питании травоядных животных зоомасса растет, но общая биомасса уменьшается, так как только часть усвояемой фитомассы превращается в зоомассу (вторичную биомассу). При питании хищников по той же причине уменьшается не только общая биомасса, но и зоомасса. По П. Дювиньо ( Duvigneaudet al., 1962) нужно 8211 кг люцерны, чтобы произвести 1035 кг телячьего мяса; последнее, будучи в свою очередь съедено детьми, дает прирост их веса только на 48 кг.
Соответственно 6,3 • 109 кал солнечной энергии дают 1,49 • 107 кал, связанных в фитомассе люцерны, 1,19 • 106 кал в зоомассе телят и 8,3 • 103 кал в теле человека (рис. 2). Анализ такого рода зависимостей позволяет сделать далеко идущие практические выводы, например вывод о будущем неизбежном возрастании в сельском хозяйстве доли растениеводства за счет животноводства и соответственно об увеличении в рационе человека растительной нищи за счет животной.
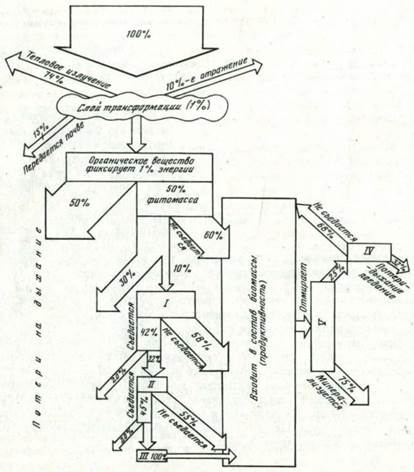
Рис. 2. Перераспределение солнечной энергии, поглощенной растительностью в ходе трофических связей на примере лесов Западной Европы, по П. Дювиньо (Duvigneaud etal., 1962) I —травоядные; II — плотоядные; III — вторичные плотоядные; IV — сапрофаги, капрофаги; V— мертвые организмы
Для изучения почвообразовательного процесса и общих закономерностей потоков вещества в биосфере представляет интерес суммарный баланс биомассы.
Может возникнуть вопрос: целесообразно ли сравнение в одинаковых единицах движения вещества и энергии в живой и неживой природе? Сравнение прихода и расхода качественно различной массы органической и неорганической материи, очевидно, смысла не имеет.
Но распределение потоков энергии, происходящей в конечном счете из одного источника, между разными процессами - биологическими, климатическими и геоморфологическими - представляет значительный интерес, в частности, для определения энергетических запасов разного вида ресурсов.
Общей единицей измерения может служить энергия, затрачиваемая или освобождающаяся в ходе отдельных процессов в единицу времени, т.е. мощность, выражаемая в киловаттах. Трактовка в калориях в час или год также возможна, но для гидрологических и геоморфологических процессов, осуществляющихся за счет превращения потенциальной энергии в кинетическую, она неудобна.
Дата добавления: 2023-11-16; просмотров: 911;
Поиск по сайту
Узнать еще
- Майнинг. Потребление электроэнергии и экология
- Обмен веществ и энергии в организме человека
- Общий тип строения центральной нервной системы. Строение серого вещества спинного мозга
- Первоначальные сведения о строении вещества
- Принцип работы гидроэлектростанции, ее мощность и выработка электроэнергии
- Строение белого вещества мозга. Нейроглия
- Строение и функции серого и белого вещества головного мозга

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
