Гиппокамп (hippocampus), или аммонов рог,
Гиппокамп (hippocampus), или аммонов рог, расположен медиобазально в глубине височных долей (см. рис. 44, 12, 46, 14; 48, 3, 8). Он имеет своеобразную изогнутую форму (гиппокамп в переводе – морской конек) и почти на всем своем протяжении образует впячивание в полость нижнего рога бокового желудочка. Гиппокамп является собственно складкой (извилиной) старой коры. С ней сращена и заворачивается над ней зубчатая извилина (рис. 47, 8).
Являясь частью старой коры, гиппокамп имеет слоистую структуру. К зубчатой извилине примыкает слой конечных ветвлений апикальных дендритов пирамидных клеток гиппокампа. Здесь они образуют молекулярный слой. На конечных разветвлениях апикальных дендритов и их основаниях оканчиваются различные афферентные волокна. Сами апикальные дендриты образуют следующий – радиальный слой. Далее, в сторону нижнего рога бокового желудочка расположен слой тел пирамидных клеток и их базальных дендритов, затем идет слой полиморфных клеток. Со стенкой бокового желудочка граничит слой белого вещества гиппокампа (alveus) (см. рис. 46, 19, 47, 3). Он состоит как из аксонов пирамидных нейронов гиппокампа (эфферентные волокна гиппокампа, уходящие в составе бахромки в свод), так и из афферентных волокон, приходящих по своду из перегородки.
Гиппокамп имеет обширные связи со многими другими структурами мозга. Он является центральной структурой лимбической системы мозга.
Лимбической системы мозга (Латинское слово limbus означает кайма, край.)
Латинское слово limbus означает кайма, край. Лимбическая система названа так потому, что корковые структуры, входящие в нее, находятся на краю неокортекса и как бы окаймляют ствол мозга. Лимбическая система включает в себя как определенные зоны коры (архипалеокортикальные и межуточные области), так и подкорковые образования (рис. 48). Из корковых структур это: гиппокамп с зубчатой извилиной (старая кора) (рис. 48, 3), поясная извилина (лимбическая кора, являющаяся межуточной) (рис. 48, 7), обонятельная кора, перегородка (древняя кора) (рис. 48, 70; из подкорковых структур: мамиллярное тело гипоталамуса (рис. 48, 7), переднее ядро таламуса, миндалевидный комплекс (рис. 48, 6).
Кроме многочисленных двусторонних связей между структурами лимбический системы существуют длинные пути в виде замкнутых кругов, по которым осуществляется циркуляция возбуждения. Большой лимбический круг (круг Пейпца) включает в себя: гиппокамп – свод – мамиллярное тело – пучок Вик д'Азира (сосцевидно-таламический пучок) – переднее ядро таламуса – кору поясной извилины – гиппокамп.
Из вышележащих структур наиболее тесные связи лимбическая система имеет с лобной корой. Свои нисходящие пути лимбическая система направляет к ретикулярной формации ствола мозга и к гипоталамусу. Через гипоталамо-гипофизарную систему лимбическая система осуществляет контроль над гуморальной системой.
Для лимбической системы характерна особая чувствительность и особая роль в ее функционировании гормонов, синтезируемых в гипоталамусе и секретируемых гипофизом, – окситоцина и вазопрессина.
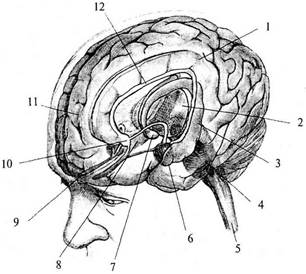
| Рис. 48. Схема строения лимбической системы: 1 – поясная извилина; 2 – таламус; 3 – левый гиппокамп; 4 – ретикулярная формация; 5 – спинной мозг; 6 – миндалина; 7 – мамиллярное тело; 8 – правый гиппокамп; 9 – обонятельная луковица; 10 – перегородка; 11 – лобная доля; 12 – свод |
Основной, целостной функцией лимбической системы является осуществление эмоционально-мотивационного поведения. Она организует и обеспечивает протекание вегетативных, соматических и психических процессов при эмоционально-мотивационной деятельности. А также осуществляет восприятие и хранение эмоционально значимой информации, выбор и реализацию адаптивных форм эмоционального поведения.
Вместе с тем каждая структура, входящая в лимбическую систему, вносит свой вклад в единый механизм, имея свои функциональные особенности. Так, функции гиппокампа связаны с памятью, обучением, формированием новых программ поведения при изменении условий. Передняя лимбическая кора обеспечивает эмоциональную выразительность речи, перегородка принимает участие в переобучении, снижает агрессивность и страх. Мамиллярные тела играют большую роль в выработке пространственных навыков, миндалевидный комплекс отвечает за пищевое и оборонительное поведение.
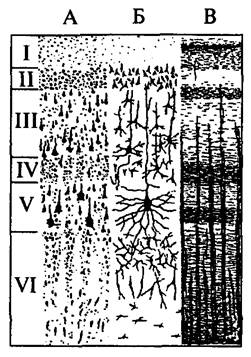
| Рис. 49. Схема цитоархитектонического строения новой коры головного мозга: А – окрашены только тела клеток; Б – окрашены отдельные клетки с отростками; В – окрашены волокна: I – молекулярный слой; II – наружный зернистый слой; III – пирамидный слой; IV – внутренний зернистый слой; V – ганглионарный слой; VI – полиморфный слой |
Новая кора.
Новая кора. Все области новой коры построены по единому принципу. Исходным типом является шестислойная кора (рис. 49). Слои представлены следующим образом:
♦ I слой – самый поверхностный, толщиной около 0,2 мм, называется молекулярным
♦ I слой – самый поверхностный, толщиной около 0,2 мм, называется молекулярным (lamina molecularis). Он состоит из волокон апикальных дендритов и аксонов, поднимающихся от клеток нижних слоев, которые контактируют друг с другом. Нейронов в молекулярном слое незначительное количество. Это мелкие горизонтальные клетки и клетки-зерна. Все отростки клеток молекулярного слоя располагаются в пределах этого же слоя.
♦ II слой – наружный зернистый (lamina granulans externa).
♦ II слой – наружный зернистый (lamina granulans externa). Толщина наружного зернистого слоя – 0,10 мм. Он состоит из мелких пирамидных и звездчатых нейронов. Аксоны этих нейронов оканчиваются на нейронах III, V и VI слоев.
♦ III слой – пирамидный (lamina pyramidalis),
♦ III слой – пирамидный (lamina pyramidalis), толщиной около 1 мм, состоит из мелких и средних пирамидных клеток. Типичный пирамидный нейрон имеет форму треугольника, вершина которого направлена вверх (рис. 50). От вершины отходит апикальный дендрит, ветвящийся в вышележащих слоях. Аксон пирамидной клетки отходит от основания клетки и направляется вниз. Дендриты клеток III слоя направляются во второй слой. Аксоны клеток III слоя оканчиваются на клетках нижележащих слоев или образуют ассоциативные волокна.
♦ IV слой – внутренний зернистый (lamina granulans internus).
♦ IV слой – внутренний зернистый (lamina granulans internus). Он состоит из звездчатых клеток, имеющих короткие отростки, и малых пирамид. Дендриты клеток IV слоя уходят в молекулярный слой коры, а их коллатерали ветвятся в своем слое. Аксоны клеток IV слоя могут подниматься в вышележащие слои или уходить в белое вещество как ассоциативные волокна. Толщина IV слоя от 0,12 до 0,3 мм.
♦ V слой – ганглионарный (lamina ganglionaris)
♦ V слой – ганглионарный (lamina ganglionaris) – слой больших пирамид. Самые крупные клетки коры расположены именно в этом слое (гигантские пирамиды Беца передней центральной извилины) (см. рис. 49Б). Их апикальные дендриты достигают молекулярного слоя, а базальные дендриты распределяются в своем слое. Аксоны клеток V слоя покидают кору и являются ассоциативными, комиссуральными или проекционными волокнами. Толщина V слоя достигает 0,5 мм.
♦ VI слой коры – полиморфный (lamina multiformis).
♦ VI слой коры – полиморфный (lamina multiformis). Содержит клетки разнообразной формы и размера, имеет толщину от 0,1 до 0,9 мм. Часть дендритов клеток этого слоя достигает молекулярного слоя, другие же остаются в пределах IV и V слоев. Аксоны клеток VI слоя могут подниматься к верхним слоям или уходить из коры в качестве коротких или длинных ассоциативных волокон (см. рис. 49).
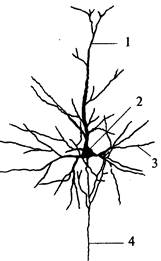
| Рис. 50. Пирамидный нейрон коры: 1 – апикальный дендрит; 2 – тело нейрона; 3 – горизонтальный дендрит; 4 – аксон |
Клетки одного слоя коры выполняют сходную функцию в обработке информации. I и IV слои являются местом ветвления ассоциативных и комиссуральных волокон, т.е. получают информацию от других корковых структур. III и IV слои являются входными, афферентными для проекционных полей, так как именно в этих слоях заканчиваются таламические волокна. V слой клеток выполняет эфферентную функцию, его аксоны несут информацию к нижележащим структурам мозга. VI слой также является выходным, но его аксоны кору не покидают, а являются ассоциативными.
Основным принципом функциональной организации коры является объединение нейронов в колонки. Колонка расположена перпендикулярно поверхности коры и охватывает все ее слои от поверхности к белому веществу. Связи между клетками одной колонки осуществляются по вертикали вдоль оси колонки. Боковые отростки клеток имеют небольшую длину. Связь между колонками соседних зон осуществляется через волокна, уходящие вглубь, а затем входящие в другую зону, т.е. короткие ассоциативные волокна. Функциональная организация коры в виде колонок обнаружена в соматосенсорной, зрительной, моторной и ассоциативной коре.
Отдельные зоны коры имеют принципиально одинаковое клеточное строение, однако есть и отличия, особенно в структуре III, IV и V слоев, которые могут распадаться на несколько подслоев. Кроме этого, существенными цитоархитектоническими признаками являются плотность расположения и размеры клеток, наличие специфических типов нейронов, расположение и направление хода миелиновых волокон (см. рис. 49В).
Цитоархитектонические особенности позволили разделить всю поверхность коры на 11 цитоархитектонических областей, включающих в себя 52 поля (по Бродману). Каждое цитоархитектоническое поле обозначено на картах мозга номером, который присваивался ему в порядке описания. Следует отметить, что между цитоархитектоническими полями не существует резких границ, клеточные слои плавно меняют свою структуру при переходе от одного поля к другому (рис. 51).
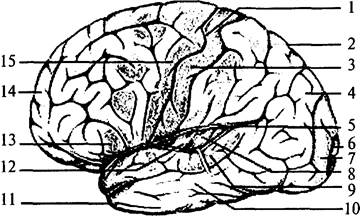
| Рис. 51. Латеральная поверхность левого полушария (Сильвиева борозда раздвинута): 1 – центральная борозда; 2 – теменная доля; 3 – задняя центральная извилина (соматосенсорная кора); 4 – ассоциативные зрительные зоны; 5 – латеральная (Сильвиева) борозда; 6 – первичное зрительное поле (участок зрительного восприятия); 7 – вторичное зрительное поле; 8 – первичный слуховой центр; 9 – вторичный слуховой центр с речевым центром Вернике; 10 – ассоциативные слуховые зоны; 11 – височная доля; 12 – островок (находится в глубине Сильвиевой борозды); 13 – двигательный речевой центр (центр Брока) – при праворукости находится только в левом полушарии; 14 – лобная доля мозга; 15 – прецентральная извилина (двигательные зоны) |
Каждое поле коры выполняет определенную функцию. Часть полей коры являются сенсорными. В первичных сенсорных полях заканчиваются проекционные афферентные волокна. Из первичных сенсорных полей информация по коротким ассоциативным волокнам передается во вторичные проекционные поля, расположенные рядом с ними. Так, поля 1 и 3, занимающие медиальную и латеральную поверхность задней центральной извилины, являются первичными проекционными полями кожной чувствительности противоположной половины поверхности тела. Расположенные рядом друг с другом участки кожи также рядом друг с другом проецируются на корковой поверхности. Такая организация проекций называется топической. В медиальной части представлены нижние конечности, а наиболее низко на латеральной части извилины расположены проекции рецепторных полей кожной поверхности головы. При этом участки поверхности тела, богато снабженные рецепторами (пальцы, губы, язык), проецируются на большую площадь коры, чем участки, имеющие меньшее количество рецепторов (бедро, спина, плечо). Поле 2, расположенное в нижнелатеральной части той же извилины, является вторичным проекционным полем кожной чувствительности.
Поля 17–19, расположенные в затылочной доле, являются зрительным центром коры, 17-е поле, занимающее сам затылочный полюс, является первичным. Прилежащие к нему 18-е и 19-е поля выполняют функцию вторичных ассоциативных полей и получают входы от 17-го поля.
В височных долях расположены слуховые проекционные поля (41, 42). Рядом с ними на границе височной, затылочной и теменной долей расположены 37-е, 39-е и 40-е, характерные только для коры головного мозга человека. У большей части людей в этих полях левого полушария расположен центр речи, отвечающий за восприятие устной и письменной речи. Поле 43, занимающее нижнюю часть задней центральной извилины, получает вкусовые афференты.
Структуры обонятельной чувствительности посылают свои сигналы в кору больших полушарий без переключений в других отделах ЦНС. Обонятельные луковицы расположены под нижней поверхностью лобных долей (см. рис. 55, 7). От них начинается обонятельный тракт, который является первой парой черепно-мозговых нервов (п. olfactorius). Корковыми проекциями обонятельной чувствительности являются структуры древней коры (рис. 52).
Двигательные области коры расположены в прецентральной извилине лобной доли (впереди от проекционных зон кожной чувствительности) (рис. 52, 1). Эту часть коры занимают поля 4 и 5. Из V слоя этих полей берет начало пирамидный путь, заканчивающийся на мотонейронах спинного мозга. Расположение и соотношение зон иннервации сходно с проекционным представительством кожного анализатора, т.е. имеет соматотопическую организацию. В медиальных частях извилины расположены колонки, регулирующие деятельность мышц ног, в нижней части, у латеральной борозды – мышц лица и головы противоположной стороны тела.
Афферентные и эфферентные проекционные зоны коры занимают относительно небольшую ее площадь. Большая часть поверхности коры занята третичными или межанализаторными зонами, называемыми ассоциативными.
Ассоциативные зоны коры занимают значительное пространство между лобной, затылочной и височной корой (60-70% новой коры). Они получают полимодальные входы от сенсорных областей коры и таламических ассоциативных ядер и имеют выходы на двигательные зоны коры. Ассоциативные зоны обеспечивают интеграцию сенсорных входов и играют существенную роль в процессах высшей нервной и психической деятельности.
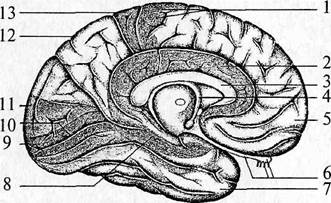
| Рис. 52. Медиальная поверхность левого полушария: 1 – прецентральная извилина (двигательные зоны); 2 – поясная извилина (часть лимбической системы), отвечает за висцеральную чувствительность; 3 – мозолистое тело (основная комиссура); 4 – свод; 5 – лобная доля; 6 – обонятельные нервы, обонятельная луковица и обонятельный путь; 7 – височная доля; 8 – гиппокамп (часть лимбической системы); 9 – первичное проекционное зрительное поле (17 поле); 10 – вторичное проекционное зрительное поле (18 поле); 11 – затылочная доля; 12 – теменная доля; 13 – задняя центральная извилина (соматосенсорные зоны) |
Филогенез.
Филогенез.У низших позвоночных (круглоротые и рыбы) развитие конечного мозга идет по пути утолщения основания, в котором формируются базальные ганглии в виде больших парных выпуклых масс. Боковые стенки и крыша конечного мозга, образующие плащ (мантию), остаются тонкими и повторяют форму полосатых тел в виде двух полушарий, хотя на два полушария конечный мозг еще не разделен, не разделена еще и полость его желудочка. В передней стенке конечного мозга развивается парное выпячивание – обонятельные доли. Они продолжаются в более или менее длинный стебелек, заканчивающийся обонятельной луковицей. Серое вещество мантии выстилает полость желудочка, а ее поверхность содержит белое вещество. В концевой пластинке располагается передняя комиссура, связывающая между собой полосатые тела.
У двоякодышащих рыб и первых наземных позвоночных (амфибии) передняя стенка мозгового пузыря вдвигается посередине между полосатыми телами в виде продольной складки, разделяющей конечный мозг на два полушария. Полость мозгового пузыря разделяется на два боковых желудочка, которые соединяются сзади Монроевыми отверстиями с III желудочком.
Мантия еще тонкая, но нервные клетки перемещаются к поверхности, однако остаются под слоем белого вещества. Над передней комиссурой появляется еще одна комиссура, соединяющая между собой мантийные структуры полушарий (аналог комиссуры гиппокампа).
У рептилий конечный мозг разрастается еще больше, но это разрастание идет в основном за счет развития базальных ганглиев. Мантия остается тонкой, но на ее поверхности впервые появляется серое вещество, которое представляет собой кору. Эта кора является высшей обонятельной структурой и сравнима с гиппокампом млекопитающих.
Усложнение организации конечного мозга млекопитающих происходит за счет развития мантии. Мантия покрыта корой. Кроме древней и старой коры в филогенетическом ряду млекопитающих появляется новая кора. Чем более высокоразвит вид млекопитающих, тем большую поверхность занимают структуры новой коры. Вместе с развитием мантии преобразуются и полости желудочков.
В добавление к имеющимся комиссурам появляется мозолистое тело, связывающее структуры новой коры обоих полушарий. С увеличением поверхности новой коры нарастает количество волокон мозолистого тела и его мощность. Мозолистое тело значительно разрастается и сдвигает назад комиссуру гиппокампа.
Кора у низших млекопитающих развивается в связи с обонятельной сенсорной системой. У высших млекопитающих (обезьяны, человека) с обонянием связаны только структуры древней и старой коры.
Кроме того, уже на ранних этапах развития позвоночных (амфибии и рептилии) кора получает афферентацию от сенсорных систем других модальностей через ядра таламуса. У млекопитающих кроме специфических ядер таламуса возникают и развиваются ассоциативные ядра, имеющие связи с неокортексом и определяющие развитие конечного мозга.
Третьим источником афферентных воздействий на кору больших полушарий является гипоталамус, который играет роль высшего регуляторного центра вегетативных функций. У млекопитающих филогенетически более древние отделы переднего гипоталамуса связаны с древней формацией коры и с древними ядрами амигдалы. Молодые отделы заднего гипоталамуса проецируются в старую кору.
Онтогенез.
Онтогенез.Вначальной стадии формирования мозга человека передний мозг является закругленным концом нервной трубки. На этом этапе конечный мозг представлен тонкой ростральной стенкой переднего мозга. Затем эта стенка выпячивается в дорсолатеральном направлении и образует два мозговых пузыря. Полости этих пузырей образуют боковые желудочки. На ростральном конце мозговой трубки и III желудочка находится участок ткани, соединяющий оба пузыря конечного мозга. В дальнейшем он станет конечной пластинкой.
Дно мешка конечного мозга утолщается очень быстро, это утолщение образует полосатое тело (стадия 10-20 мм). В дальнейшем оно делится на хвостатое ядро, скорлупу, бледный шар и миндалину. По мере роста полушарий базальные ядра смещаются медиально и примерно на 10-й неделе развития (40 мм) сливаются с промежуточным мозгом.
Растущие аксоны покидают кору примерно на 8-й неделе развития (стадия зародыша – 23 мм). Волокна, которые не заканчиваются в полосатом теле, идут вдоль плоскости слияния основания конечного мозга с промежуточным, образуя внутреннюю капсулу. Волокна, идущие каудально и огибающие снизу средний мозг, образуют ножки мозга. Затем на вентральной поверхности продолговатого мозга они образуют пирамиды. Уходя в спинной мозг, пирамидные пути перекрещиваются и оканчиваются на мотонейронах спинного мозга. Ассоциативные проводящие пути начинают выявляться в конце второго месяца развития. Свод появляется из гиппокампа в конце третьего месяца развития.
Мозолистое тело появляется в начале четвертого месяца развития. Оно начинает формироваться в концевой пластинке в виде пучка поперечных волокон, лежащих над комиссурой гиппокампа. Мозолистое тело растет очень быстро в каудальном направлении и смещает туда же комиссуру гиппокампа и свод. Гиппокамп также смещается назад.
Пузыри конечного мозга разрастаются в ростральном, дорсальном и каудальном направлениях. На ранних стадиях развития (до 3–4-го месяца) пузыри имеют очень тонкую стенку и утолщаются медленно.
Борозды и извилины начинают формироваться только с 11 – 12-й недели. Первыми появляются латеральная и гиппокампова извилины. Затем формирование борозд протекает очень быстро. К моменту рождения существуют все основные извилины.
ОРГАНЫ ЧУВСТВ
В процессе эволюции у животных сформировались системы восприятия окружающего мира – экстерорецепторы – и системы оценки состояния собственных систем организма – интерорецепторы.
Анатомически органы чувств состоят из воспринимающей части – совокупности рецепторов, проводящих нервных путей, промежуточных центров обработки информации и коркового отдела, в котором происходит распознавание стимулов.
Разные по своему строению и внутренней организации рецепторы позволяют организму воспринимать различные по своим свойствам внешние и внутренние сигналы. К ним относятся: световые, звуковые, обонятельные, тактильные, вкусовые, температурные, механические, болевые и другие сигналы или раздражители.
Рецепторный компонент органов чувств состоит из специальных клеток, имеющих специфическое строение и способных воспринимать определенный тип сигнала. Проводниковый компонент представлен нервными волокнами, идущими к соответствующим структурам центральной нервной системы – спинному и головному мозгу, где и происходит обработка непрерывно поступающей информации.
Зрительная система
Орган зрения состоит из глазного яблока, расположенного в глазнице, и зрительного нерва, идущего к соответствующим участкам коры головного мозга
Ранее уже упоминалось, что в процессе эмбриогенеза архэнцефалона происходит превращение его в передний мозговой пузырь, из дорсолатеральных стенок которого выступают два глазных пузыря. В дальнейшем из них развиваются некоторые компоненты зрительной системы, в том числе проводящие пути промежуточного мозга.
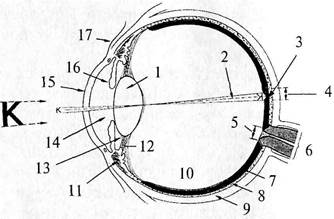
| Рис. 53. Схема горизонтального сечения правого глаза: 1 – хрусталик; 2 – зрительная ось; 3 – центральная ямка; 4 – желтое пятно; 5 – диск зрительного нерва; 6 – зрительный нерв; 7 – сетчатка; 8 – сосудистая оболочка; 9 – склера; 10 – стекловидное тело; 11 – ресничная мышца; 12 – волокна пояска; 13 – задняя камера; 14 – передняя камера; 15 – роговица; 16 – радужная оболочка; 17 – конъюнктива |
Глазное яблоко
Глазное яблоко имеет шаровидную форму, которая изменяется в процессе постнатального развития новорожденного. Оно состоит из ядра, покрытого тремя оболочками – фиброзной, сосудистой и сетчатой (внутренней) (рис. 53).
Фиброзная оболочка подразделяется на прозрачную переднюю часть – роговицу (рис. 53, 15) и заднюю – склеру (рис. 53, 9). Склера (sclera) представляет собой плотную соединительную ткань, образованную пучками коллагеновых волокон. Сзади на склере находится решетчатая пластинка, через которую проходят волокна зрительного нерва. В толще склеры, на границе соединения ее с роговицей, имеется сеть мелких полостей, образующих венозный синус склеры, через который происходит отток жидкости из передней камеры глаза.
Роговица (cornea) – это выпуклая пластинка блюдцеобразной формы
Роговица (cornea) – это выпуклая пластинка блюдцеобразной формы, круглый край (лимб) которой переходит в склеру. Толщина роговицы от 0,8 до 1, 1 мм. Роговица лишена кровеносных сосудов, и ее питание происходит за счет лимфы.
Сосудистая оболочка глазного яблока находится под склерой и состоит из собственно сосудистой оболочки, ресничного тела и радужки. Ресничное тело участвует в аккомодации глаза, поддерживая, фиксируя и растягивая хрусталик. Большая часть ресничного тела – это ресничная мышца (рис. 53, 11), образованная пучками миоцитов, среди которых различают продольные, циркулярные и радиальные волокна.
Ресничное тело спереди продолжается в радужку (рис. 53, 16), которая представляет собой круглый диск с отверстием в центре (зрачок). Радужка, в свою очередь, состоит из пяти слоев. В толще одного из них (сосудистого) проходят две мышцы, пучки миоцитов которых образуют сфинктер (сжиматель) зрачка и радиально расположенные пучки, расширяющие зрачок (дилататор зрачка). Расширительные пучки иннервируются постганглионарными симпатическими волокнами клеток, лежащих в верхнем шейном узле; сжиматель – постганглионарными парасимпатическими волокнами ресничного узла. Пигментный слой радужки двухслойный, а цвет зависит от количества меланина.
Хрусталик
Хрусталик (рис. 53, 1), представляет собой прозрачную двояковыпуклую линзу, диаметром около 9 мм, имеющую переднюю и заднюю поверхности, переходящие одна в другую в районе экватора. Ось хрусталика – линия, соединяющая наиболее выпуклые точки обеих поверхностей, имеет размеры от 3, 7 до 4, 4 мм. Хрусталик покрыт прозрачной капсулой. Ядро хрусталика образовано прозрачными волокнами призматической формы, состоящими из белка кристаллина. Эти волокна формируются в эмбриональном периоде и сохраняются в течение всей жизни.
Хрусталик подвешен на ресничном пояске (цинновой связке) (рис. 53, 72), между волокнами которого расположены петитов канал, сообщающийся камерами глаза. Волокна пояска в сочетании с мышцами обеспечивают аккомодацию глаза.
Стекловидное тело (рис. 53, 10) представляет собой аморфное межклеточное вещество, на передней поверхности которого в ямке расположен хрусталик.
Две камеры глаза, в которых циркулирует влага, содержащая около 0,02% белка, создают внутреннюю среду глазного яблока.
Внутренняя оболочка глазного яблока – сетчатка (retina) прилежит изнутри к сосудистой оболочке. Она состоит из двух листков – внутреннего светочувствительного (нервная часть) и наружного пигментного. Различают заднюю зрительную (фоторецепторную) и переднюю, не содержащую фоторецепторов, части.
Сетчатка имеет слоистое строение (рис. 54). Наружный – пигментный слой (рис. 54, 7), состоит из пигментных эпителиоцитов, от внутренней поверхности которых отходят отростки, отделяющие друг от друга палочки и колбочки (фоторецепторы). Пигментный слой поглощает световые лучи, предотвращая их отражение. К пигментному эпителию прилежит слой палочек и колбочек, которые представляют собой периферические отростки фоторецепторов. Каждый фоторецептор состоит из наружного и внутреннего сегментов. Наружный – светочувствительный сегмент имеет впячивания плазматических мембран. У палочек эти мембраны образуют диски, у колбочек – мембранные складки. В мембране наружных сегментов содержатся зрительные пигменты. Внутренний сегмент фоторецепторов содержит митохондрии, рибосомы и другие элементы клетки.
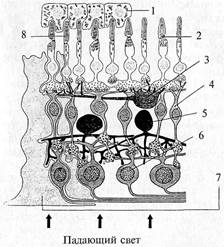
| Рис. 54. Структура сетчатки: 1 – пигментный эпителий; 2 – колбочка; 3 – горизонтальная клетка; 4 – биполярная клетка; 5 – амакриновая клетка; 6 – ганглиозная клетка; 7 – мюллерова клетка; 8 – палочка |
Основой фоторецепции является реакция распада пигментов под действием света. Палочки содержат пигмент родопсин и отвечают за сумеречное (черно-белое) зрение. Колбочки различаются по содержащимся в них пигментам на три типа. Один из них содержит пигмент, реагирующий на воздействие красного диапазона световых лучей, другой имеет пигмент, разлагающийся под действием зеленого цвета, а третий реагирует на синий диапазон спектpa. Таким образом, полный набор всех трех типов колбочек обеспечивает цветное зрение.
В сетчатке глаза человека насчитывается около 6–7 млн колбочек и от 70 до 120 млн палочек. В центре сетчатки находится углубление – центральная ямка (см. рис. 53, 3), в которой очень плотно расположены колбочки. По наличию окраски это место называется еще желтым пятном. При помощи глазных мышц изображение рассматриваемого объекта проецируется в область центральной ямки, что позволяет лучше различить детали. Изображение, проецируемое за пределы центральной ямки, попадает в поле периферического зрения.
Ребенок рождается с развитым черно-белым восприятием окружающего мира, цветное зрение развивается уже в постнатальный период.
От каждой фоторецепторной клетки отходит отросток, который образует синапс с отростками биполярных клеток II слоя (рис. 54, 4). Биполярные клетки выполняют функцию усилителя сигнала и передают информацию ганглиозным клеткам сетчатки (рис. 54, 6), которые являются выходными элементами сетчатки, так как их аксоны (500 тыс. – 1 млн) образуют зрительный нерв.
Кроме перечисленных клеток в сетчатке имеются клетки, участвующие в регуляции функционирования других клеток. Это горизонтальные и амакриновые клетки.
Сетчатка является не только местом восприятия, но и первичным нервным центром обработки зрительной информации.
Место выхода из сетчатки зрительного нерва называется диском зрительного нерва (слепое пятно) (рис. 53, 5). В центре диска в сетчатку входит центральная артерия сетчатки.
Зрительные нервы проникают в полость черепа через каналы зрительных нервов (рис. 55, 56). На нижней поверхности мозга образуется перекрест зрительных нервов – хиазма (рис. 56, 6), причем перекрещиваются только волокна, идущие от медиальных частей сетчаток. После перекреста зрительные пути называются трактами. Большинство волокон зрительного тракта направляются в латеральное коленчатое тело промежуточного мозга (рис. 56, 5). Латеральное коленчатое тело имеет слоистое строение и названо так потому, что его слои изгибаются наподобие колена. Нейроны данной структуры направляют свои аксоны через внутреннюю капсулу, затем в составе зрительной радиации к клеткам затылочной доли коры больших полушарий возле шпорной борозды. Этот путь является специфическим зрительным, по нему идет информация только о зрительных стимулах.
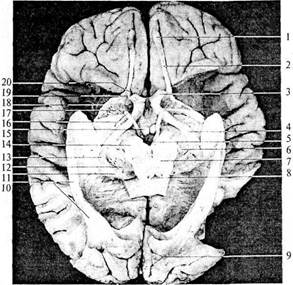
| Рис. 55. Препарат вентральной поверхности мозга (височные доли и ствол сняты, средний мозг рассечен): 1 – обонятельная луковица; 2 – обонятельный тракт; 3 – зрительный нерв; 4 – зрительный тракт; 5 – межножковая ямка; 6 – блоковый нерв; 7 – черная субстанция среднего мозга; 8 – водопровод мозга; 9 – борозда птичьей шпоры; 10 – зрительный пучок; 11 – подушка таламуса; 12 – медиальное коленчатое тело; 13 – латеральное коленчатое тело; 14 – ножка среднего мозга; 15 – сосцевидное тело; 16 – глазодвигательный нерв; 17 – переднее продырявленное пространство; 18 – латеральная обонятельная полоска; 19 – обонятельный треугольник; 20 – медиальная обонятельная полоска |
Другая часть волокон, направляясь к верхним буграм четверохолмия (рис. 56, 9), составляет неспецифический зрительный путь. От клеток верхнего холмика аксоны идут к ядру глазодвигательного нерва (рис. 55, 76), который иннервирует мышцы глаза и зрачка, замыкая, таким образом, рефлекторные дуги быстрых реакций на зрительные стимулы.
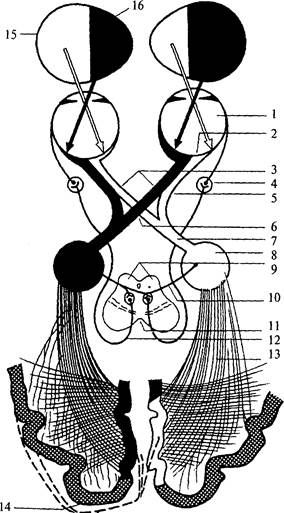
| Рис. 56. Схема проводящих путей зрительного анализатора: 1 – глазное яблоко; 2 – сетчатка; 3 – зрительные нервы; 4 – ресничный ганглий; 5 – неперекрещивающиеся волокна зрительного нерва; 6 – хиазма; 7 – зрительный тракт; 8 – латеральное коленчатое тело; 9 – верхние бугры четверохолмия; 10 – неспецифический зрительный путь; 11 – средний мозг; 12 – волокна парасимпатической нервной системы; 13 – зрительная радиация; 14 – зрительная кора; 15 – латеральное поле зрения; 16 – медиальное поле зрения |
Через ручки верхних холмиков четверохолмия волокна неспецифического зрительного пути направляются к ядрам подушки таламуса, затем в зрительную кору (рис. 56, 14).
Зрительная кора располагается взатылочных долях больших полушарий и занимает 17–19-е поля, по Бродману.
Теперь рассмотрим вспомогательные структуры глаза: мышцы век и слезный аппарат.
Различают шесть глазодвигательных мышц – четыре прямые, две косые. Пять из шести мышц начинаются в глубине глазницы в окружности зрительного канала от общего сухожильного кольца, окружающего зрительный нерв и глазную артерию. Мышцы сокращаются и расслабляются согласованно, благодаря чему оба глазных яблока двигаются синхронно.
К комплексу анатомических структур, составляющих орган зрения, относятся также веки и слезный аппарат. Веки защищают глазное яблоко спереди. Они образуют своими кожными складками структуру, ограничивающую глазную щель, а в медиальном углу образуют «слезное озеро», на дне которого виден рудимент третьего века. Веки снабжены соответствующим мышечным аппаратом и сальными железами. По краям век расположены ресницы, выполняющие определенную защитную функцию. Защитную роль в структуре глаза играет и конъюнктива, служащая переходом от век к глазному яблоку.
Слезные железы, имеющие слезные протоки, которые открываются в верхний свод конъюнктивы, выполняют смачивающую и защитную функции. Последняя обеспечивается за счет наличия в составе слезы лизоцима, обладающего выраженным антибактерицидным действием.
Развитие органа зрения в филогенезе претерпело ряд сложных эволюционных этапов – от одиночных светочувствительных клеток до глаза млекопитающих, обладающих цветным бинокулярным зрением.
Органы слуха
Слуховой аппарат включает в себя три отдела: наружное ухо, среднее и внутреннее.
Наружное ухо
Наружное ухо состоит из хрящевой ушной раковины и наружного слухового прохода, расположенного в височной кости и выстланного серными железами. Отделяется наружное ухо от среднего барабанной перепонкой (рис. 57, 12).
Среднее ухо
Среднее ухо представляет собой полость, ограниченную с одной стороны барабанной перепонкой, а с другой – структурами внутреннего уха (рис. 57Б). Полость среднего уха называется барабанной и выстлана слизистой оболочкой. При помощи Евстахиевой трубы (рис. 57, 77) полость сообщается с носоглоткой. В среднем ухе находится система косточек: молоточек, наковальня и стремечко (рис. 57М, Н, С), которые являются усилителями звуковой волны. Они передают волновые колебания от мембраны барабанной перепонки к мембране овального окошка внутреннего уха. На внутренней стенке среднего уха имеется два отверстия (окна) – овальное (рис. 57, 3) и круглое (рис. 57, 5). И круглое, и овальное окна закрыты мембраной, причем к мембране овального окошка прикреплено стремечко среднего уха.
Внутреннее ухо
Внутреннее ухо – полое костное образование в височной кости, разделенное на костные и перепончатые каналы и полости, содержащие рецепторный аппарат слухового и статокинетического (вестибулярного) анализаторов (рис. 57В). Внутреннее ухо находится в толще височной кости и состоит из системы сообщающихся друг с другом костных каналов – костного лабиринта, в котором расположен перепончатый лабиринт. Очертания перепончатого лабиринта почти полностью повторяют очертания костного. Пространство между костным и перепончатым лабиринтом, называемое перилимфатическим, заполнено жидкостью – перилимфой, которая по составу сходна с цереброспинальной жидкостью. Перепончатый лабиринт погружен в перилимфу, он прикреплен к стенкам костного футляра соединительнотканными тяжами и заполнен жидкостью – эндолимфой, по составу неско
Дата добавления: 2016-06-05; просмотров: 5646;
Поиск по сайту
Узнать еще
- Base-5 или толстый Ethenet
- CIH — газораспределительный механизм с распределительным валом в головке блока цилиндров
- Cемейство Мотыльковые (Fabaceae, или Leguminosae)
- Cравнительная характеристика усилителей на БТ
- D-технология построения чертежа. Типовые объемные тела: призма, цилиндр, конус, сфера, тор, клин. Построение тел выдавливанием и вращением. Разрезы, сечения.
- II. Организация дезинфекционных и стерилизационных мероприятий в организациях, осуществляющих медицинскую деятельность
- IV.Стабилизация нефти.
- MRP мы уже определили.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
