Разнообразие клеток
Методы исследования клетки
Цитология – наука о микроскопической и субмикроскопической структуре клетки и ее жизнедеятельности.
Клетка – это элементарная структурная и функциональная единица тела растений и животных, способная к самовоспроизведению. Первым увидел клетку английский естествоиспытатель Р. Гук при изучении покровной ткани бузины – пробки. Он усовершенствовал микроскоп, изобретенный Г. Галилеем в 1609 г., и использовал его для исследования тонких срезов органов растений. Свои наблюдения Р. Гук изложил в сочинении «Микрография», изданном в 1665 г., где он впервые применил термин «клетка». В 1833 г. английский ботаник Р. Броун обнаружил ядро, а в 1839 г. чешский физиолог Я. Пуркинье – цитоплазму. Многочисленные наблюдения клеточного строения растений и животных позволили немецким ученым – ботанику М. Шлейдену и зоологу Т. Шванну – в 1838-1839 гг. сформулировать клеточную теорию, суть которой заключается в том, что клетка – это основная элементарная структурная единица всех живых организмов. Клеточная теория доказывает единство происхождения, строения и эволюции растений и животных.
К концу XIX в. цитология окончательно сформировалась в самостоятельную науку. На базе светового микроскопа были изучены основные компоненты клетки, накапливались данные об их функциях.
Дальнейший прогресс цитологии связан с изобретением электронного микроскопа. В биологии его начали использовать в середине XX в.
Применяемые для изучения клеток методы очень разнообразны. Основной из них – микроскопический. Большую роль продолжает играть световой микроскоп, современные модели которого дают увеличение до 2 тыс. раз. Но возможности светового микроскопа ограничены, частицы менее 0,2 мкм рассмотреть при помощи такого микроскопа невозможно. Электронный микроскоп дает увеличение в 200-300 тыс. раз и более. Здесь вместо пучка света используют поток электронов, движущихся с высокой скоростью.
Методом культуры тканей изучают структуру и жизнедеятельность живых клеток вне организма.
Цитохимический метод позволяет выявить наличие и определить количество различных веществ в клетке.
Разделить компоненты клетки с различной плотностью для изолированного изучения их можно с помощью метода центрифугирования. Извлечь из клетки отдельные компоненты (ядро, митохондрии и др.) позволяет метод микроскопической хирургии.
Строение клетки
Разнообразие клеток
Все разнообразие форм клеток можно свести к двум группам:
- паренхимные клетки – длина равна ширине или превышает ее не более чем в 2-3 раза;
- прозенхимные клетки – длина превышает ширину во много раз.
Средняя длина клеток высших растений 10-100 мкм. Наиболее крупные паренхимные клетки достигают в длину несколько миллиметров и видны невооруженным глазом, например клетки плодов арбуза, лимона, клубней картофеля. Но особенно большую длину имеют прозенхимные клетки стеблей льна (40 мм), крапивы (80 мм), рами (200 мм).
Компоненты клетки
Рассматривая взрослую растительную клетку при помощи светового микроскопа, можно увидеть следующие компоненты: плотную стенку, одну большую или 2-3 небольшие вакуоли, занимающие центральную часть клетки; цитоплазму, расположенную между стенкой клетки и вакуолью; находящееся в цитоплазме ядро.
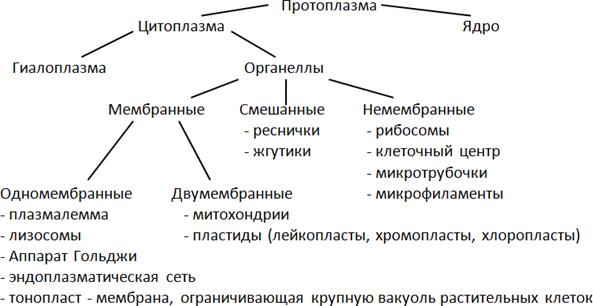
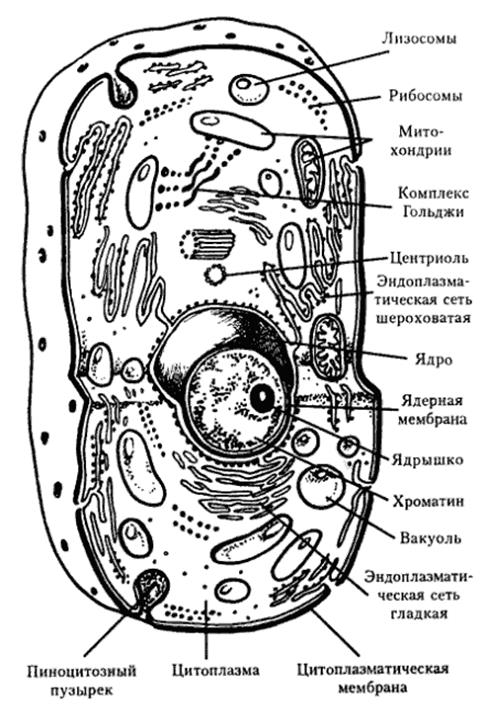
Рис. 1. Структурные компоненты эукариотической клетки
Протопласт (протоплазма) – содержимое растительной клетки, кроме клеточной оболочки.
Ядро и цитоплазма – живые части клетки и в совокупности составляют протопласт.
Стенка и вакуоли – неживые части клетки, производные протопласта, продукты его жизнедеятельности.
Цитоплазма– сложная структурная система, имеет мембранную организацию, состоит из гиалоплпзмы и органелл. Ее структуру образуют тонкие (4-10 нм), довольно плотные пленки – биологические мембраны. Основу их составляют липиды. Молекулы липидов расположены упорядоченно – перпендикулярно к поверхности, в два слоя, так, что их части, интенсивно взаимодействующие с водой (гидрофильные), направлены наружу, а части, инертные по отношению к воде (гидрофобные), – внутрь. Молекулы белка расположены несплошным слоем на поверхности липидного каркаса с обеих его сторон. Часть их погружена в липидный слой, а некоторые проходят через него насквозь, образуя участки, проницаемые для воды.
Мембраны образуют пограничный слой цитоплазмы, а также внешнюю границу ее органелл и участвуют в создании их внутренней структуры. Они делят цитоплазму на изолированные отсеки, в которых одновременно и независимо друг от друга могут протекать биохимические процессы часто в противоположном направлении (например, синтез и распад).
Одно из основных свойств биологических мембран – избирательная проницаемость (полупроницаемость): одни вещества проходят через них с трудом, другие легко и даже в сторону большей концентрации. Мембраны во многом определяют химический состав цитоплазмы и клеточного сока.
Плазмодесмы – тончайшие цитоплазматические нити, которые осуществляют связь между клетками. Клеточная пластинка, образующаяся при цитокинезе, пронизана трубочками эндоплазматического ретикулума, которые не разобщаются. На их основе и формируются плазмодесмы. Стенка плазмодесменного канала выстлана плазмалеммой, соединяющей плазмалеммы смежных клеток. В центре канала проходит трубка, сохраняющая непрерывность ретикулума обеих клеток. Между плазмалеммой и трубкой находится гиалоплазма, также непрерывная для обеих клеток. Плазмодесмы чаще всего бывают собраны в группы по нескольку десятков. Поодиночке они располагаются в стенках, не имеющих вторичных утолщений.
Плазмалемма и тонопласт. Плазмалемма – это мембрана, отграничивающая цитоплазму от стенки клетки и обычно плотно прилегающая к ней. Иногда плазмалемма бывает волнистой или образует глубокие складки. Она регулирует обмен веществ клетки с окружающей средой, а также участвует в синтезе веществ. Тонопласт отграничивает цитоплазму от вакуоли. Функция его та же, что и плазмалеммы.
Гиалоплазма. Это жидкая непрерывная среда, в которую погружены органеллы. Гиалоплазма содержит ферменты и нуклеиновые кислоты. Белки, входящие в состав гиалоплазмы, образуют сеть из тонких фибрилл (диаметром 2-3 нм) – трабекулярную систему, которая связывает между собой органеллы. Эта система очень динамична, она может распадаться при изменении внешних условий. Гиалоплазма способна к активному движению, которое может быть вращательным – вдоль стенки клетки, если в центре находится одна большая вакуоль, и струйчатым – по тяжам, пересекающим центральную вакуоль, в разных направлениях. Скорость движения зависит от температуры, интенсивности света, снабжения кислородом и других факторов. При движении гиалоплазма увлекает за собой органеллы. Гиалоплазма осуществляет взаимосвязь органелл, участвует в обмене, транспорте веществ, передаче раздражения и т. д.
Эндоплазматический ретикулум (эндоплазматическая сеть). Представляет собой отграниченную, мембранами систему взаимосвязанных субмикроскопических каналов и цистерн, пронизывающих гиалоплазму. Имеются две формы ретикулума: гранулярный (шероховатый) и агранулярный (гладкий). Гранулярный эндоплазматический ретикулум несет на поверхности мелкие гранулы – рибосомы. Агранулярный эндоплазматический ретикулум состоит из ветвящихся трубочек, отходящих от цистерн гранулярного ретикулума, не имеет рибосом. Обычно он развит слабее, чем гранулярный. Участвует в синтезе и транспорте эфирных масел, смол, каучука.
Рибосомы. Это гранулы диаметром около 20 нм, расположенные в гиалоплазме или прикрепленные к поверхности мембран эндоплазматического ретикулума. Они обнаружены также в митохондриях и пластидах. Рибосомы состоят из белка и рибонуклеиновой кислоты (РНК) и не имеют мембранной структуры. Функция рибосом – синтез белка, самовоспроизводство живой материи. Этот процесс происходит в рибосомах, расположенных группой и связанных между собой нитевидной молекулой и-РНК. Такие группы называют полисомами. Считают, что рибосомы формируются в ядре.
Аппарат Гольджи. Состоит из диктиосомы и пузырьков Гольджи. Диктиосома представляет собой стопку из 5-7 плоских цистерн, ограниченных агранулярной мембраной. Диаметр цистерн около 1 мкм, толщина 20-40 нм. Цистерны не соприкасаются друг с другом. Пузырьки Гольджи отчленяются от краев цистерн и распространяются по всей гиалоплазме. В диктиосоме происходят синтез, накопление и выделение полисахаридов. Пузырьки Гольджи транспортируют их, в том числе и к плазмалемме. Мембрана пузырьков встраивается в плазмалемму, а содержимое оказывается снаружи от плазмалеммы и может включаться в стенку. Пузырьки Гольджи могут включаться и в тонопласт.
Происхождение диктиосом еще точно не установлено. Считают, что в их образовании принимает участие эндоплазматический ретикулум. В некоторых клетках аппарат Гольджи отсутствует.
Сферосомы. Округлые блестящие тельца диаметром 0,5-1 мкм. Это центры синтеза и накопления растительных масел. Они отшнуровываются от концов тяжей эндоплазматического ретикулума. Мембрана, расположенная на поверхности сферосомы, по мере накопления масла редуцируется, и от нее остается только наружный слой.
Лизосомы. Пузырьки размером 0,5-2 мкм, имеющие на поверхности мембрану. Содержат ферменты, которые могут расщеплять белки, липиды, полисахариды и другие органические соединения. Образуются так же, как и сферосомы, из тяжей эндоплазматического ретикулума. Их функция – разрушение отдельных органелл или участков цитоплазмы (локальный автолиз), необходимое для обновления клетки.
Митохондрии. Форма митохондрий чрезвычайно разнообразна – овальная, округлая, цилиндрическая, гантелевидная, ветвистая и т.д. Длина их равна 2-5 мкм, диаметр – 0,3-1 мкм. На поверхности митохондрии находятся две мембраны. Внутренняя мембрана образует выросты в полость митохондрии в виде гребней или трубочек, называемых кристами. Кристы значительно увеличивают мембранную поверхность митохондрии. Пространство между кристами заполнено жидким веществом – матриксом, в котором находятся рибосомы и содержится дезоксирибонуклеиновая кислота (ДНК). Поверхность внутренней мембраны покрыта мельчайшими тельцами, имеющими шаровидную головку и ножку (АТФ-сомы).
Митохондрии – это энергетические лаборатории клетки. Здесь происходят расщепление углеводов, жиров и других органических веществ при участии кислорода (дыхание) и синтез АТФ. Выделяемая при дыхании энергия преобразуется в энергию макроэргических связей молекулы АТФ, которая затем используется для осуществления процессов жизнедеятельности клетки – деления, поглощения и выделения веществ, синтеза и т.д.
Пластиды. Бывают только у растений. Эти органеллы имеют на поверхности две мембраны. В зависимости от окраски различают три типа пластид:
- хлоропласт – зеленого цвета;
- хромопласты – желтого, оранжевого, красного цветов;
- лейкопласты – бесцветные.
Хлоропласты содержат зеленый пигмент хлорофилл, а также пигменты из группы каротиноидов – каротин (оранжевый) и ксантофилл (желтый). Именно с хлорофиллом связана основная функция хлоропластов – синтез органических веществ из неорганических при участии энергии света (фотосинтез). Поэтому хлоропласты присутствуют только в клетках наземных органов, на которые падает солнечный свет. Хлоропластам растения обязаны зеленой окраской.
У высших растений хлоропласты в большинстве случаев имеют линзовидную форму. Диаметр их 4-6 мкм, толщина 1-3 мкм. У водорослей хлоропласты, называемые также хроматофорами, очень разнообразны по форме и величине. Они могут иметь звездчатую форму, лентовидную, сетчатую и др. Обычно в клетке имеются от 1 до 50 хлоропластов. Располагаются они в постенном слое цитоплазмы.
Внутри хлоропластов находится однородное вещество – строма, пронизанная системой параллельно расположенных мембран. Мембраны имеют вид плоских мешков, их называют тилакоидами, или ламеллами. У большинства высших растений часть тилакоидов имеет дисковидную форму. Эти тилакоиды собраны в стопки, называемые гранами. Хлорофилл и каротиноиды находятся в каждой из двух мембран тилакоида граны. Граны связаны между собой тилакоидами стромы. Внутренняя мембрана оболочки хлоропласта иногда образует складки и переходит в тилакоиды стромы. В строме находятся молекулы ДНК, рибосомы, капли липидов, называемые пластоглобулами, крахмальные зерна и другие включения.
Крахмал образуется в хлоропластах из продуктов фотосинтеза, его называют фотосинтетическим, или первичным. При помощи ферментов фотосинтетический крахмал осахаривается и в виде глюкозы транспортируется из листа на построение органов или в запас.
Лейкопласты не содержат пигментов. По размеру они значительно меньше хлоропластов и не имеют строго определенной формы. Лейкопласты присутствуют чаще всего в клетках тканей и органов, на которые не падает солнечный свет: в корнях, клубнях, семенах и др. Внутренняя мембранная система у лейкопластов развита значительно слабее, чем у хлоропластов. В строме имеются молекулы ДНК, рибосомы, пластоглобулы. Основная функция лейкопластов – синтез и накопление запасных питательных продуктов, в первую очередь крахмала, иногда белков, редко масла. Лейкопласты, накапливающие крахмал, называют амилопластами. В них из сахаров, поступающих из фотосинтезирующих органов, образуются крахмальные зерна различного размера и формы - вторичный крахмал. Запасной белок может откладываться в виде кристаллов или аморфных гранул, масло - в виде пластоглобул.
Хромопластысодержат красные, оранжевые, желтые пигменты из группы каротиноидов. По размеру они меньше хлоропластов, форма их очень разнообразна. Внутренняя мембранная система у них чаще всего отсутствует.
В зависимости от формы накопления каротиноидов различают хромопласты глобулярного, фибриллярного (трубчатого) и кристаллического типов. У хромопластов наиболее распространенного глобулярного типа пигменты растворены в пластоглобулах. У хромопластов фибриллярного типа каротиноиды не только присутствуют в пластоглобулах, но и образуют группы параллельных нитей или трубок, расположенных в строме. У хромопластов кристаллического типа каротиноиды присутствуют главным образом в виде кристаллов различной формы, определяющих форму самой пластиды (серповидную, ромбовидную, игловидную и т. д.).
Хромопласты встречают в клетках лепестков некоторых растений, зрелых плодов, осенних листьев. Их функция в процессе обмена веществ не выяснена. Косвенное биологическое значение хромопластов состоит в привлечении насекомых для перекрестного опыления и животных для распространения семян.
В процессе эволюции первыми из пластид появились хлоропласты, из которых при расчленении тела растения на органы образовались лейкопласты и хромопласты. В онтогенезе почти все виды пластид могут переходить друг в друга. Наиболее часто происходит превращение лейкопластов в хлоропласты (например, при образовании зародыша из оплодотворенной яйцеклетки) и хлоропластов в хромопласты (например, при осеннем пожелтении листьев). Лишь хромопласты в природных условиях, как правило, не превращаются в другие виды пластид. Количество пластид в клетке увеличивается за счет деления их путем перетяжки.
Дата добавления: 2018-05-10; просмотров: 5257;
Поиск по сайту
Узнать еще
- Автоматическая работа и взаимозависимость клеток организма
- В митотическом цикле клеток
- В-система иммунитета – это система органов, клеток и эффекторных молекул, осуществляющих гуморальную форму иммунного реагирования.
- ВАКУОЛИ РАСТИТЕЛЬНЫХ КЛЕТОК
- ВЗАИМОДЕЙСТВИЕ КЛЕТОК ПРИ КЛЕТОЧНОМ ИММУННОМ ОТВЕТЕ.
- ВИДЫ ПОВРЕЖДЕНИЙ И ГИБЕЛИ КЛЕТОК. УНИВЕРСАЛЬНЫЙ ОТВЕТ КЛЕТКИ НА ПОВРЕЖДЕНИЕ
- Визуальное разнообразие
- Вирусы – фрагменты наших клеток

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
