Экологические формы
Вертикальная С и н у з и и
ярусность
Жизненные формы Пространственные
Горизонтальная
мозаичность Сезонные
Фитоценотипы
Экотопогенная К о н с о р ц и и
Фитогеографические Фитогенная Консоргенты Консорты
и флорогенические
элементы Антропогенная Индивидуаль- Растения
Ные
Животные
Популяцион-
Ные
Синузиальные
Рис. 7.3. Характерные свойства фитоценозов
Выделяются по отношению к внешним факторам среды характерные группы растений (табл. 7.2).
Таблица 7.2
Основные наземные экологические группы растений по отношению к разным факторам среды (по О.Е. Агаханьянц, 1968)
| Экологический фактор | Экологическая группа растений при интенсивности воздействия фактора | ||
| недостаточной | умеренной | сильной | |
| Тепло | Микротермы | Мезотермы | Макротермы |
| Влага | Ксерофиты | Мезофиты | Гигрофиты |
| Минеральное | Олиготрофы | Мезотрофы | Эвтрофы |
| питание | |||
| Засоленность | Олигогалофиты | Мезогалофиты | Эвгалофиты |
| почв | |||
| Песчаный грунт | Псаммофиты | ||
| Обломочный грунт | Хасмофиты | ||
| Скальный грунт | Литофиты | ||
| Холодные сухие условия | Криофиты | ||
| Холодные влажные условия | Психрофиты | ||
| Солнечное освещение | Гелиофиты | ||
| Ветер | Сциофиты | Теневыносливые | Анамофиты |
Ценоэлементы - термин введенный Х..Х. Трассом. Это пространственно ограниченные участки фитоценоза, отличающиеся по составу и внешнему виду. Описание ценоэлементов предполагает выделение синморфологических и синэкологических единиц. К синморфологическим факторам относится ярусность и горизонтальная мозаичность. Вертикальный ярус растительности играет эдификаторную роль в формировании расположенных под ним ценоэлементов. Под пологом верхнего яруса условия существования растений значительно трансформированы.
Ценоэлементы горизонтальной структуры фитоценозов называются микрогруппировками или микроценозами. В растительных сообществах различают экотопогенную, антропогенную и фитогенную мозаичность. Первая связана с неровностями микрорельефа или неоднородностью почвогрунтов в пределах одного экотона. Благодаря мозаичности полнее используется разнообразие условий микросреды, увеличивается устойчивость фитоценоза.
К синэкологическим элементам относятся синузии и консорции.
Синузии образованы группами видов, подчас далеких в систематизационном отношении, но сходных по внешнему виду. Например, деревья, травы и кустарнички, мхи и лишайники в таежном лесу. По форме смен травяных растений можно судить о стадиях дернового процесса. В местах срубленного леса распространены ползучекорневищные злаки (костер, пырей, полевица белая, вейник и др.), среди которых есть корнеотпрыски (осоты, жирухи и др.), бурьяны (крупные зонтичные, крапивы и др.). Это первая так называемая корневищная стадия дернового процесса.
С уплотнением почвы они сменяются рыхлодерновинными - тимофеевкой, овсяницей луговой, ежой сборной и др. Вместе разрастаются бобовые - клевера, мышинный горошек, люцерна. Это наиболее хозяйственно ценная стадия развития.
Третяя - плотнодерновинная, плотнокустовая стадия, когда разрастаются овсяница овечья, щучка, белоус.
Сезонные синузии представлены разными биоформами, развивающимися на одной территории в разные сроки вегетационного периода, то есть отграничены друг от друга во времени. Например: синузия эфемеров в пустыне.
Чем больше синузий в сообществе, тем больше его флористическое, экологическое и биоморфное богатство.
У растительности есть и свои специфические способы связывания соседних фитоценозов: встречная экспансия популяций "сшивает" растительный покров в единую ткань (континиум).
Связующим звеном между фито- и зооценозом является консорция. Например, сосна и ее микоризмы грибки, с эпофитными мхами и лишайниками на стволах и ветвях, с паразитическими грибками, со множеством населяющих ее членистоногих.
Консорция состоит из консоргента (ядра консорции) и консортов - группы организмов, связанных с ядром. Консоргент играет роль эдификатора. Это автотроф. Типичные консорты ведут малоподвижный образ жизни. Разнообразие консортов - свидетельство зрелости сообщества, отсутствие говорит о молодости биоценоза.
Каждый ландшафт характеризуется своим спектром экологических рядов.
Важнейшими экологическими аспектами, связанными с состоянием современных ландшафтов и их биогеографическими особенностями, являются проблемы обезлесения и опустынивания.
Обезлесение
Леса - один из первоисточиков жизни. Это регулятор кислорода биомассы, своеобразная "легочная" система планеты. Климатоохранные функции леса сравнимы с океаническими. Водоохранные функции лесов отражаются в зависимости величины поверхностного стока атмосферных вод от степени залесенности. Величина подобного стока для безлесной местности составляет 65% от суммы годовых осадков, а для лесной зоны всего 5%. Залесенность сохраняет почвы. Если в лесостепных условиях слой почвы в 18 см смывается за 65 лет, то в лесу за многие тысячелетия.
При обезлесении ухудшается циркуляция подземных вод. Особенно ярко это проявилось в Индии, где проводятся интенсивные рубки леса в Гималаях. Из-за этого только с 1960 по 1980 гг. площадь ежегодных наводнений здесь удвоилась. К 1985 г. затапливалось уже до 59 млн га, вместо 19 млн га в 1960 г. Такая же картина наблюдается и в Бангладеш, где затопляемые в результате наводнений площади доходят до 2/3 территории страны.
Особенно ценны экваториальные леса: они наиболее древние, крупнопородные и переполнены разнообразными живыми организмами. Здесь сосредоточено 50% всех видов растений и животных, несмотря на то, что они занимают всего 6% площади суши.
Лесов с сомкнутыми кронами насчитывается в мире 28 млн км2, общая площадь сплошных и разреженных лесов составляет менее трети от размеров суши свободной ото льдов. Естественные и саженные леса покрывают 35 млн км2.
В результате антропогенной деятельности было уничтожено не менее 10 млн км2 лесов. Весьма показательны в этом плане сравнительные схемы сокращения лесов в Европе и США (рис. 7.4, 7.5).

Рис. 7.4. Облесение центральной Европы с начала X по начало XX в., по I.G. Simmons
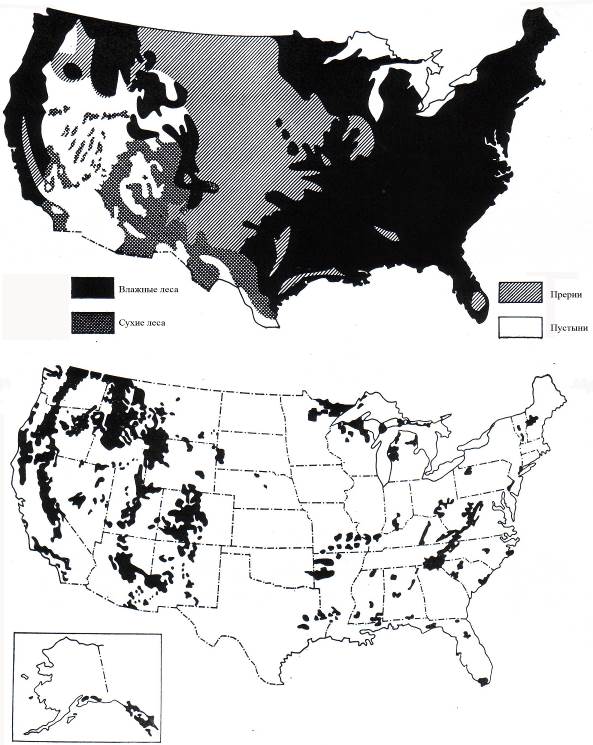
Рис. 7.5. Уничтожение лесов в США (Dasman, 1972)
Первоначальное распространение лесов (вверху); современные леса (внизу)
Современные темпы потери лесов оцениваются от 11 до 20 млн га в год. А 17 млн га - это площадь равная территории Австрии.
Немецкий географ А. Гумбольд высказался образно и горько:
"Человеку предшествуют леса, а его сопровождают пустыни" (рис. 7.6).
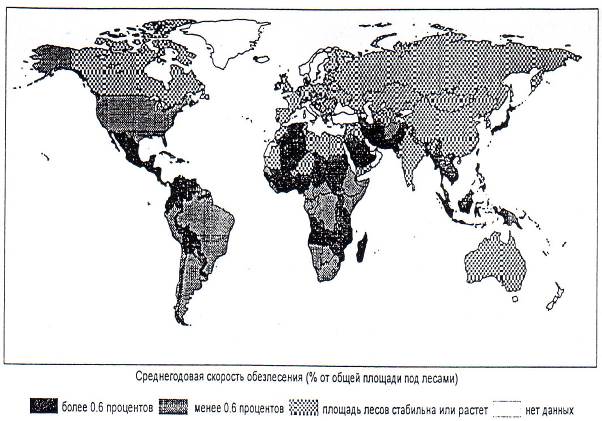
Рис. 7.6. Средняя величина обезлесения за год по странам мира, в %% от общей площади лесов за период с 1980 по 1990 гг.
За двадцать лет (с 1970 по 1990 гг.) мир потерял дополнительно почти 200 млн га лесных массивов (это примерно площадь такой страны как США, восточнее Миссисипи).
Однако, не следует бросаться и в крайности. Ряд исследователей полагали, что при нынешних темпах вырубки влажных тропических лесов они могут полностью исчезнуть после 2010-2020 гг. Но анализ данных по Амазонии и данные спутниковых съемок последнего 20-летия показывают, что общая площадь потерь здесь лесов за почти 30 лет составила примерно 5 млн км2 (порядка 6% от исходной площади леса этого региона). Причина этого очевидно в том, что рубкам в основном подвергаются территории экотонной полосы расположенной между лесными и травянными формациями. Наиболее опасны для сокращения лесов строительно-дорожные работы, приводящие к фрагментации и островизации самих лесных экосистем.
Самое важное, что всегда следует иметь в виду, - это то, что экосистемная функция лесов, в том числе и особенно тропических, незаменима.
Опустынивание
Площади засушливых земель на нашей планете составляют уже 6,1 млрд. га. За год опустыниванию подвергается 60 тыс. км2 (2 Бельгии или в 1,5 раз больше территории Республики Алтай). На грани опустынивания находится сейчас в мире 20 млн км2 земель (вдвое больше чем площадь Канады). Становится все больше пустынь и опустыненных земель. Только за 20-летие (1970-1990 гг.) пустыни увеличились на 120 млн га (это площадь пахотных угодий в Китае). Засушливые земли занимают уже 1/3 суши (рис. 7.6).
В соответствии с климатическими особенностями пустыни могли бы составить площадь около 48 млн км2 (включая ледяные). Фактически их площадь сейчас 57 млн км2. Следовательно, порядка 9 млн км2 - это антро-
погенные пустыни. Опустынивание еще развивается на площади 25 млн км2. Одна шестая населения мира живет в условиях угрозы опустынивания (рис. 7.7).
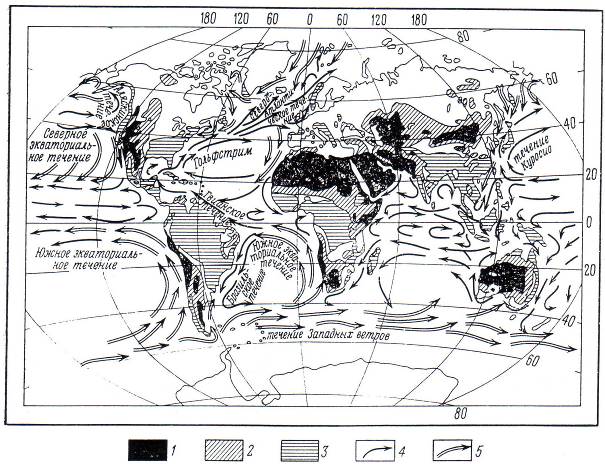
Рис.7.7. Мировая карта современных пустынь, аридных зон, холодных и теплых океанических течений и областей с количеством годовых осадков более 1000 мм/год
Черным показаны пустыни, диагональной штриховкой – области с годовым количеством осадков 10-100 мм, белым – области с годовым количеством осадков 100-1000 мм, горизонтальной штриховкой – области с годовым количеством осадков более 1000 мм .
1 – аридные области; 2 – полуаридные области; 3 – осадков более 1000 мм/год; 4 – теплое течение; 5 – холодное течение
Характерно, что лишь немногие из территорий с очень высокой степенью опустынивания (около 3 млн км2, или в 6 раз больше чем Франция) примыкают к абсолютным пустыням, хотя и расположены не в зоне пустынь, а полупустынь. В связи с этим, наряду с мнением, что пустыня наступает на наименее плодородные территории, существует и другая, не менее обоснованная: парадокс, но симптоматичный - самую большую нагрузку земля испытывает там, где лучше условия. Иными словами, пустыни не столько разрастаются, сколько создаются человеком. И с этих пози-ций крайне показательно сопоставление последовательности сельскохозяйственного освоения земель в мире с современным размещением самих пустынь.
Очень выразительны развития событий в зоне Сахеля. Это обширная территория к югу от пустыни Сахары. В 1968 г. здесь началась длительная (20 лет) засуха (рис. 7.8).
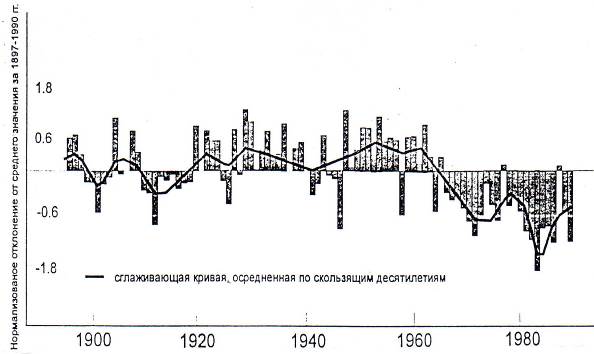
Рис. 7.8. Колебания годовых сумм осадков в Сахеле за
1897-1990 гг.
В результате ее первой волны (1968-1978 гг.) от голода погибло более 250 тыс. жителей и 40% скота (в Мали и Мавритании свыше 90%).
Любопытно, что здесь в скотоводческой зоне производительность пастбищ в 1,5-10 раз выше, чем в современных хозяйствах Австралии и Техаса (используются все виды растительности, вплоть до ветвей деревьев). Практикуется почти исключительно отгонный метод скотоводства: зимой скот перегоняют к югу, летом - на север.
Кроме того, здесь и численность населения превышает потенциальную емкость территории, а местами это сочетается с противоречивыми интересами скотоводов и земледельцев. С этих позиций Сахель обычно подразделяется на земледельческую и животноводческую зоны, примерно по изогете 400 мм атмосферных осадков.
Вместе с тем, последствия опустынивания здесь оказались одни из самых грозных. Эффективная борьба с опустыниванием должна опираться на глубоком проникновении в экологические проблемы этого массового явления.
3. Биопродуктивность
Объем первичной биологической продукции определяется общим количеством органического вещества образованного на определенной площади за единицу времени (обычно год). "Чистая" биопродуктивность, кроме того, учитывает расход синтезированного органического вещества на дыхание растений. Эти величины выражаются через массу сухого органического вещества, или содержащегося в нем углерода. Отсюда средний коэффициент пересчета сухой массы на углерод составляет 0,45, обратный - 2,2.
Для суши Земли биопродуктивность за год составляет 130 млрд. т оргвещества, или около 60 млрд.т С; для Мирового океана - 90 и 40 млрд. т, соответственно. То есть общая среднегодовая продуктивность равна 220 млрд. т органического вещества, или 100 млрд. т углерода. Удельная продуктивность для Мира 430 г/м2 (43 ц/га), в том числе для суши (без ледниковых областей) 1000 г/м2, для океана 250 г/м2.
В основном это фитомасса и главным образом леса (87%).
Общая масса живого вещества Земли оценивается в 1300 млрд. т органического вещества (590 млрд. т С), по Д. Алькамо (1994).
Баланс углерода для первичных экосистем при длительности их функционирования до 1000 лет квазистационарен. Естественная эволюция экосистем при этих условиях определяется приростом в 0,1% от биопродуктивности. Этот остаточный член баланса носит название экосистемной продуктивности.
В результате антропогенной деятельности в настоящее время в атмосферу выносится 1,6 7+ 01,0 млрд. т С/год, что составляет 3% (!) биопродукции. К 2050 г. чистая первичная продукция достигнет величины 82,5 млрд. т/год, при экосистемной продукции 8,1 млрд. т.
Таким образом, степень разбалансированности увеличится до 10%.
Глобальный биогеохимический цикл углерода по сути - основа стабилизирующей роли биоты. Согласно В.Г. Горшкову (1995), синтез и деструкция органического вещества на глобальном уровне балансируются с точностью 10-4 для диапазонов в 10000 лет.
Первичная биопродуктивность естественных ландшафтов определяет верхний предел продуктивности непреобразованных систем. Антропогенные воздействия ведут к ее снижению. В мире 15 млн км2 земли уже занято под земледелие и дают 2,5 млрд. т сельхозпродуктов, определяя среднюю урожайность порядка 17 ц/га.
Весьма значительна роль биоты в глобальном гидрологическом цикле. Живое вещество на 90% состоит из воды. Тогда биота связывает во вновь формирующемся органическом веществе около 500 км3 воды. Во время фотосинтеза растительность пропускает через себя на два порядка воды больше, чем оказывается связанной в самом органическом веществе. Поэтому в биологическом звене глобального круговорота воды участвует порядка 30000 км3 воды в год. А это уже почти 25% суммарных осадков, выпадающих на сушу.
Эмпирически установлено, что изменение энергетики экосистем на 1% выводит ее из стационарного состояния. Не случайно на фотосинтез приходится только 0,16% приходящей суммарной солнечной радиации.
В.Г. Горшковым рассчитано, что человек должен использовать не более 1% чистой первичной биопродукции, чтобы оставить за биотой контролирующие функции по устойчивости развития биосферы. 99% в этом случае идет на поддержание ее устойчивости.
По законам биологической экологии человеку на питание полагается не более 10 млрд. т биопродукции в год. Сейчас он уже использует 31 млрд. т. Да вследствие антропогенных изменений первичной продукции она сократилась на 27 млрд. т. Следовательно, общее количество потребляемой и разрушаемой человеком биомассы достигло 58 млрд. т/год, или почти 40% первичной биологической продукции. Подобные же расчетные величины получены и П. Витусеком с соавторами (США).
Этим же автором разработана концепция о биотической регуляции окружающей среды. Согласно ей два важнейших эмпирических фактора не находят объяснения при традиционной существующей концепции: почему, несмотря на быстрые изменения окружающей среды, особенно происходящие под воздействием самой биоты, условия окружающей среды не выходят за пределы возможности существования любой жизни, и почему, несмотря на непрерывную адаптацию, все виды сохраняют строгую дискретность и не наблюдается переходных форм ни между современными, одновременно существующими в биосфере, видами, ни видами, наблюдаемыми по палеоданным. В.Г. Горшковым биота Земли рассматривается как единственный механизм поддержания пригодных для жизни условий окружающей среды в локальных и глобальных масштабах.
Взаимодействие с окружающей средой приводит к необходимости образования сообществ видов и экологических систем определенного набора видов, скоррелировано взаимодействующих между собой и окружающей их средой. Тогда виды, поддерживающие данные условия окружающей среды, очевидно, должны сохранять неизменной свою генетическую программу и не могут непрерывно адаптироваться к любым флуктуациям внешних условий.
Все виды биоты должны быть не только жизнеспособны, но и скоррелировано взаимодействовать с другими видами сообщества, обеспечивая управление окружающей средой.
Эволюционные переходы к новым видам ограничены и происходят только при сохранении способности к стабилизации окружающей среды в составе экологического сообщества.
В.Г. Горшков идею Г. Лавлока о гомеостазе глобальной экосистемы объяснил подчиненностью биосферы принципу Ле-Шателье - Брауна, в связи с чем она ведет себя как саморегулирующаяся система. Основные выводы, дополненные впоследствии, таковы: биота Земли способна с высочайшей точностью поддерживать пригодное для жизни состояние окружающей среды; огромная мощность продукции, достигнутая ею, позволяет восстанавливать любые естественные нарушения окружающей среды в краткие сроки - десятки лет; огромная мощность, накопленная ей, таит скрытую опасность такого же быстрого разрушения глобальной экосистемы, если ее целостность будет нарушена; биосфера, вероятно, может компенсировать те нарушения, производимые человеком, пока доля его потребления не превысит 1% продукции биосферы; современные антропогенные изменения биосферы, с выбросом биотой 2,3 млрд.т/год углерода в атмосферу, показывают на ее неустойчивое состояние; это состояние еще обратимо, но следует на порядок снизить потребление ее естественной продукции; какого либо другого устойчивого состояния биосферы не существует; необходимо снизить численность людей на планете почти на порядок и только тогда дестабилизированная биосфера способна будет возвратиться к состоянию ее устойчивости, так как отторжение ее продукции человечеством не будет превышать 1%.
Дата добавления: 2021-11-16; просмотров: 531;
Поиск по сайту
Узнать еще
- III. Суммарные допуски формы расположения
- V. Сборка и нагружение формы.
- V.VII. Зависимость ширины и формы выхода слоя на поверхности от его истинной мощности, угла падения и формы рельефа
- VI 1.4. Геоэкологические проблемы земледелия
- VII. Геоэкологические проблемы использования почвенных и земельных ресурсов
- А - с прямолинейной спинкой; б - с криволинейной спинкой; в - с канавкой удлиненной формы
- А) Контроль отклонений от правильной цилиндрической формы.
- А. Локализованные формы

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
