Морская подсистема биосферы
Как известно, в океанических просторах по глубинам и морфологии дна выделяются площади шельфа - 8 (7,6-8,4%) морского дна, континентального склона (15,3%) и самого океанического ложа.
По разделению условий глубинности жизнеобитания шельф отвечает литоральной, склон - батиальной, ложе - абиссальной зонам.
В связи с подобным районированием океанических областей, для самого Мирового океана характерным является значительное различие экологических условий жизнеобитания. Биоцикл океана и моря распадается на два биохора (пространства, занятых группами схожих биотопов): донную - бентальную область (бентос) и пелагиальную область открытого моря - пелагиаль.
Собственно морские биоценозы делятся на бенталь и пелагиаль.
Выделяются пассивно плавающие животные и растения - планктон (зоо- и фито-) и активно плавающие - нектон. Пелагель делится на неритическую - прибрежную и океаническую части.
Морская подсистема биосферы занимает центральное положение в ее структурно-функциональной организации. Оценивать значимость биосферных блоков следует по количеству участвующей солнечной энергии. Последние оценки: величина первичной продукции фотосинтеза в океане ок. 100×109т С/год-1 (100 гигатонн), в то время как продукция растений суши составляет 51×109 т С/год-1.
Биомеханические циклы биосферы замыкаются через Мировой океан. Практически вся океанская вода - биокосное тело. За 2 млн лет она полностью проходит через биотический круговорот.
Лишь в биоразнообразии биосфера океана уступает наземной. Особенно большое разнообразие биотопов связано с огромной массой крупной наземной растительности, а от этого зависит существование массы экологических ниш. Фантастически разнообразен класс насекомых (1,5 млн видов). Наоборот, пищевые цепи океана, особенно пелагиали, базируются на мельчайших одноклеточных водорослях фитопланктона и сравнительно однотипны. Благодаря подвижности биотопа (воды), океан более гомогенен и число экологических ниш не может быть большим, откуда и снижение биоразнообразия. Исключение - пограничные области (вода-дно, вода-суша). Но биоразнообразие суши частично определено океанами, как непреодолимыми преградами, изолирующими фации и флоры островов и материков.
Структура экосистем океана имеет принципиальное отличие от экосистем суши.
Биокосное вещество на суше - тонкий слой почв. В океане - это вся толща вод.
Только этим двум природным телам - почве и воде – присущи свойства, которые обеспечивают процессы продуцирования. На суше это 1 м, в океане 4000 м. В океане плодородие верхнего продуцирующего слоя (30-150 м) обеспечивается вертикальным перемешиванием за счет запасов глубинных вод, в почвах общий запас биогенных веществ лишь немного превышает их годовой расход или равен ему.
Основная особенность общества пелагиали - создаваемое фитопланктоном первичное органическое вещество продуцируется в слое до 100-150 м. Животное население только этого тонкого слоя постоянно обитает вместе с растительным эргоценом. Население остальной громады вод живет за счет органического вещества продуцируемого в поверхностной зоне. Характерна разобщенность глубин первичного продуцирования органического вещества и значительной части его потребления. Эта-то разобщенность и определяет основу процессов трансформации вещества и циклов отдельных элементов в толще вод океана.
Таким образом, если на суше оставшиеся массы консументов и редуцентов обитают совместно с продуцентами, то в океане основная масса консументов отделена от продуцентов и основное направление эволюции преследует выработку приспособлений к возможно более полному использованию органики, идущей сверху.
Может быть и в этом одна из причин сравнительно низкого видового разнообразия океана. Около 0,1% величины первичной продукции захоранивается в осадках и выходит надолго из глобального круговорота.
Захоронение углерода в океане происходит по действием "биологического насоса":
1.Переход минерально растворенного углерода в виде СО2-ионов в минерально нерастворимые СаСО3 (менее MgCO3), идет главным образом в водах океана тропических областей. А на глубине свыше 4000 м происходит полное растворение этих минералов (лизоклин).
2. Еще более важным является включение СО2 во вновь образуемые в результате фотосинтеза органические вещества. При объеме годовой продукции 100×109 т С это составляет 37×1010 т СО2.
Распределение первичной продукции фотосинтеза в океане приведено на рис. 7.1.
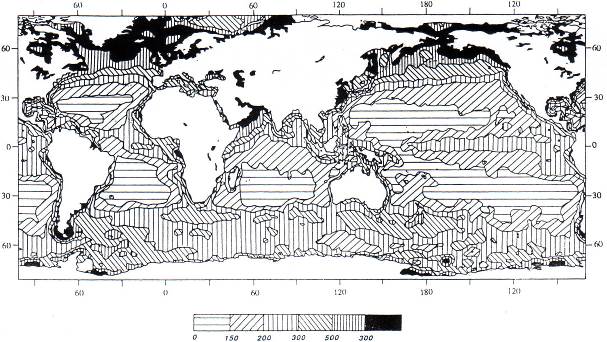
Рис. 7.1. Распределение первичной продукции фотосинтеза в океане
Фотосинтез
В среднем для всего океана превышение поглощения СО2 над выделением в атмосферу за год составляет 2×109 т (некоторые полагают 9 Гт).
Величина захоронения углерода на суше заметно отличается лишь в тундре и тайге. Так что, "именно северные болота и тайга, а не тропические леса, являются долговременными поглатителями углерода и, следовательно, продуцентами кислорода, то есть "легкими планеты". В океане же, на всей его громадной площади преобладает поглощение углерода из атмосферы, хотя в обширных олиготрофных районах эта величина очень низка" (с. 113).
Основой жизнеобеспечения на Земле является, главным образом, фотосинтез. Он представляет собой "изумительно совершенный" механизм преобразования световой энергии в сахара (углеводы), которые являются основой питания всех живых организмов. Под воздействием энергии коротковолновой части солнечных лучей хлорофилл – сложное соединение С, Н, Mg и N - адсорбирует и разлагает молекулы воды. При этом водород, соединяясь с углеродом и другими химическими элементами, образует углеводы, а кислород выделяется в атмосферу. В синтезе участвуют и небольшие количества минеральных веществ почвы. Этот процесс значительно совершеннее чем технологии любого современного промышленного производства. Поэтому все происходит без отходов и загрязнений. Последующие процессы биосинтеза приводят к образованию протеинов и липидов.
Процессу фотосинтеза противоположен идущий одновременно процесс дыхания: выделение углекислого газа и образование в качестве конечных продуктов воды и полезной энергии. Таким образом, вода и углерод - вещества, связанные в начале процесса фотосинтеза, в конце его высвобождаются, а кислород, выделившийся прежде, теперь вновь адсорбируется.
От правильности функционирования этих двух механизмов - круговорота кислорода, вкупе с меньшими по объемам круговоротами азота, серы и фосфора,- зависит на Земле вся дыхательная жизнь растений и животных.
Природа действует экономно.
6 СО2 + 6 Н2О
днем ночью
(солнечная (теплоотдача
энергия) энергии)
С2Н12О6 + 6 О2
(глюкоза)
Вместе с тем, не надо преувеличивать и КПД фотосинтезирующей деятельности (отношение расхода энергии на синтез биомассы по отношению к общему количеству приходящей солнечной энергии), который обычно составляет 0,1-1%.
Также "неэкономно" растительный покров расходует и водные ресурсы. Продуктивность транспирации (отношение прироста веса сухой массы растений к расходу воды на транспирацию за определенный промежуток времени) обычно составляет от 1/200 до 1/1000 (чаще 1/300). Но это и естественно, поскольку транспирация представляет собой механизм замедленного, но равномерного тока привноса вещества путем диффузии в виде интенсивно разбавленных растворов.
Орган ассимиляции фотосинтезирующего растения - лист - представляет собой футляр из плотной катикулярной ткани, пронизанной массой мелких отверстий - устьиц, которые открываются и закрываются. В футляре находится большая поверхность хлоропластов, содержащих зерна хлорофилла. Для процесса фотосинтеза с ассимиляцией углекислоты необходима влажная растворяющая среда. Диффузия углекислоты сопровождается обратной диффузией водяного пара. Поэтому растения обычно и используют небольшую часть (порядка 10%) возможного диффузионного притока углекислоты. При усвоении 1 г углекислоты растение в среднем теряет путем транспирации около 360 г воды.
Величина солнечной энергии, используемой для построения органики в процессе фотосинтеза на Земле, составляет 133×1012 ватт.
Это больше общемирового уровня потребления электроэнергии в 13 раз, но по отношению к приходящей к поверхности Земли солнечной энергии составляет всего 0,16%. Отношение затрат энергии на фотосинтез к поглощению солнечной энергии составляет от 0,1 до 1% (в среднем 0,5%).
Передача энергии от первичной биологической продукции к более высоким уровням экологической пирамиды сопровождается ее потерями. Отношение биомассы организмов к количеству потребляемого ими органического вещества обычно не превышает 10-20%. В природных системах с одного трофического уровня на другой, более высокий переходит в среднем не более 10%.
Вероятно, это один из аппаратов устойчивости жизни при значительных диапазонах колебаний в поставках энергии и вещества.
В обычных условиях энергия фотосинтеза медленно отдается в атмосферу, практически почти не влияя на метеорологические процессы. Но это весьма заметно при крупных пожарах, когда энергия фотосинтеза быстро превращается в тепло и метеорологические условия быстро меняются.
Хемогенез
В последние годы выясняется, что существуют неизвестные и мало известные нам органические миры. Возможно, что своеобразные биосферы присутствуют на отдельных планетах Солнечной системы. Таким образом, вероятно, следует говорить о системах биосфер.
Свыше 110 лет назад русский микробиолог С.Н. Виноградский открыл хемосинтез - образование некоторыми бактериями органических веществ из двуокиси углерода за счет энергии, получаемой путем окисления восстановленных неорганических соединений. Как известно, обычно за единственный процесс формирования органических веществ считался фотосинтез, идущий с использованием солнечной энергии. Исследования дна океана за последние десятилетия (4-5) позволили выделить в многочисленных зонах спрединга и субдукции литосферных плит, зонах теплового разложения газогидратов и т.п. до глубин свыше 6 км множество участков, населенных специфическими сообществами донных организмов, существующих на основе хемосинтеза.
Следовательно, все население Мирового океана по происхождениюделится на фотобиос и хемобиос. Сообщества хемобиоза всегда крайне ограничены по площади. Но биомасса здесь вместо обычных граммов достигает 40-60 кг/м2, то есть по масштабам и насыщенности органики не уступают даже самым богатым мидиевым банкам и устричным формациям на мелководье умеренных широт. Уже описано более 15 новых семейств и более 160 7новых видов организмов. Известны очень крупные экземпляры. Длина погонофор (рифтии) достигает 1,5 м. Размеры двустворчатых моллюсков достигает 25-30 см в поперечнике, толщина раковины – около 8 мм. Растут они в 500 раз быстрее, чем их ближайшие родственники в «обычных» условиях. Исследование этой фауны сулит нам немало сенсационных открытий.
Л.Лобье предполагает наличие на дне моря подповерхностных полостей, где могут быть обнаружены новые для науки живые организмы.
Близится к завершению еще один грандиозный проект, предпринятый более четверти века назад. Обсуждается вопрос о завершающей стадии бурения мощной толщи льда в районе антарктической станции "Восток", где вероятно существование подледного озера. К зиме 1996 г. глубина скважины достигла 3350 м и вскрыла льды возраста около 420 тыс. лет. Керн этой скважины содержит данные об истории глобальных изменений климата Земли на протяжении последних четырех ледниково-межледниковых циклов.
Мощность ледникового покрова здесь равна 3700-4200 м. В 1970-х гг. с помощью радиозондирования здесь выявлены крупные скопления подледных вод. Уже полученные результаты свидетельствуют о следующей прогнозной характеристике озера Восток. Его местоположение 76 50 030' ю.ш., 102-106 50 0 в.д. Длина его 230 км, ширина 50 км, площадь поверхности около 10 тыс. км2. Глубина до 500 м, возможная мощность осадков на дне 100-200 м. Температура поверхностного слоя воды -2,4 до
-3,4°С.
Это совершенно новое для нас природное явление, которое является географическим открытием мирового значения.
Озеро было изолировано от внешнего мира в течении миллиона и более лет. Из льда, таящего на верхней кромке озера должен поступать атмосферный воздух, попавший туда сотни тысяч и миллионы лет назад. В этих условиях, очевидно, способна существовать микрофауна и микрофлора, представляющие исключительный научный интерес.
Уже существуют доказательства присутствия здесь органической жизни: на больших глубинах во льду обнаружены микроорганизмы.
"Биосистемы воспроизводятся и развиваются только за счет ресурсов планеты и под действием космических сил - питательные вещества, энергия взаимодействия, перетоки информации... Биологические системы вбирают в себя из среды не любое, а только необходимое вещество - кислород и водород воздуха, воду и водород гидросферы, макро- и микроэлементы из почвенных растворов и минералов. Формируясь на базовом для них субстрате, они адаптируются к экосистемам нормативного в каждой географической зоне хемотипа".
В экологическом аспекте хемотипы экосферы должны иметь исключительное значение по связям с химическими параметрами среды обитания. "Еще Э. Геккель, введший термин экология, относил к взаимосвязи организмов и среды все условия их существования, которые не сводятся единственно к биологическому их содержанию. Так, невозможно рассматривать условия жизни без обращения к явлениям фотохимии в атмосфере, биохимии в биосфере, геохимии и гидрохимии в лито- и гидросфере и биохимии в педосфере. От всех них зависит химическая активность и функционирование живого вещества, выраженное в процессах обмена".
Высказывается мнение, что, кроме хемосинтеза, в качестве энергии для биоценоза, возможны и другие нетрадиционные источники. К этому подводит новая концепция организованности подземной биосферы Земли, согласно которой граница биосферы в литосфере может достигать глубины в 7 и более км, и состоит из последовательных зон бактериальных фильтров: верхнего – углеводородокисляющего и нижнего – углекисло-водородного (рис. 7.2).
Эти «фильтры» выполняют, на наш взгляд, роль аналогичную «биологическому насосу» в морских обстановках. Геохимическое значение этой деятельности отражено в табл. 7.1.
Возможно, что это биоценозы, которые могут быть обусловлены просто явлениями термосинтеза.
В зоне активного водообмена происходит природный процесс микробиологического оксигенеза – образования молекулярного кислорода, прямо или косвенно связанный с жизнедеятельностью алканотрофных бактерий.
Ниже зоны окислительного бактериального фильтра в восстановительной обстановке затрудненного водообмена находится зона нафтидо биоза. По мнению А.А. Оборина, нафтидобиоз – современный процесс микробиологического синтеза газообразных и жидких углеводородов природным биоценозом подземных вод.
Фотосинтез
СО2+Н2О=Сорг+О2↑
_____________________ ö_________________________________
öСН4+О2→СО2+Н2О
Зона углеродокисляющего
бактериального фильтра
_______________________________________________________
Зона бактериального нафтидобиоза ÷
СН4 ö на орг. в-ве фотосинтеза
Метаногенез
Сорг СО2 СО НСО3- на орг. в-ве хемосинтеза
________________________________________________________
ù
Н2+СО2→СН4∙СН3СООН∙Сорг
Зона углекисло-водородного
бактериального фильтра
________________________________________________________
öН2 СО СО2ö
Рис. 7.2. Схема функционирования подземной биосферы
2. Устойчивость биосферы
Существует понятие гомеостаза - совокупности сложных приспособительных реакций человека и животного, направленных на устранение или максимальное ограничение действия различных факторов внешней и внутренней среды, нарушающих относительное постоянство внутренней среды организма (например, постоянство температуры тела, содержания глюкозы в крови). Понятие гомеостаза было введено американским физиологом У. Конноном (Cannon, 1932). Позднее оно было перенесено на природные процессы саморегуляции биосферы, на экологические системы.
Таблица 7.1
Схема геохимической деятельности биоценоза подземной гидросферы
| Исходные компоненты | Микроорганизмы | Продукты жизнедеятельности (экзометаболиты) |
| Аэробные | ||
| CH4, N2 | Метанотрофы | CO2+H2O, N+орг, Сорг, жирн. кислоты – С18, лизин, метионин, треонин |
| С3H8-C4H10, N2 | Пропан-бутанотрофы | CO2+H2O, N+орг, Сорг, аргинин, валин, лизин, метионин, треонин, ПАВ |
| жидкие у/в, N2 | Нефтеокисляющие бактерии, актиномицеты, водоросли Bacillus mycoides и др. | CO2+H2O, Сорг, ПАВ, полисахариды |
| СO2 H2S, [FeS, CuS, MoS2], CO2 | Тионовые бактерии, Thiobacillus ferrooxidans | CaCO3 |
| H2S, CO2 | Серобактерии | H2SO4, сульфаты, [Fe2(SO4)3], |
| N2, CO2, О.В. | Азотфиксаторы, цианобактерии | N2 S |
| CO2, NH4+ | Нитрифицирующие литотрофы | NH4+-NH3, Nорг, H2 |
| CO2, C1-соед., NH4+ | Метилотрофы | NO3-, Cорг |
| NH4+, О.В. | Гетеротрофы | NO3-, Cорг |
| Орг. кислоты, спирты, NO3 | Гетеротрофные денитрификаторы | NO3-, Cорг |
| CO2, NO3-, О.В. | Литотрофные денитрификаторы | H2O, полиакриламид (CH2=CH-CO-NH2) |
| CO2, H2 | Водородные бактерии | N2,Cорг H2O, Cорг |
| Анаэробные | ||
| H2, CO, CO2, О.В. CO2, SO4, О.В. | Метаногены | CH4, Cорг, Nорг, B12 (Co-содержащие), изопреноидные у/в –C30, C25, C20 |
| О.В. | Сульфатвосстанавлива-ющие бактерии | H2S, сульфиды (FeS), CH4, у/в C8-C24, Nорг, CO2-CaCO3, меркаптаны |
| NO3-, NO2-, О.В. | Водородобразующие бактерии | Н2 |
| H2, CO, О.в. | Денитрифицирующие бактерии Ацетогены | N2, CO2 CH3COOH |
Биосфера возникла одновременно с появлением жизни, то есть для Земли - несколько млрд. лет назад. Все компоненты биосферы обладают ограниченной устойчивостью. И, вместе с тем, сохранение жизни на Земле в течении такого длительного времени вероятно только при сравнительно малой изменчивости окружающей среды в целом.
Поскольку экологические системы (как и отдельные организмы) - объекты длительного эволюционного развития, то в его процессе, вероятно, сохранялись лишь те экологические системы, устойчивость которых по отношению к диапазону колебаний внешних факторов была достаточно значительной.
Ограниченная устойчивость компонентов при малой изменчивости системы возможна только в хорошо отлаженной комплексной системе, включающей в себя согласованную иерархию систем различной степени сложности. Они должны быть взаимосвязаны, чтобы результатирующий общий эффект всегда был достаточно равновесным.
Так, устойчивость климата Земли определяется обратной отрицательной связью зависимости уходящего в космос длинноволнового излучения от температуры воздуха у земной поверхности. Колебания уровней водоемов на наклонных берегах зависят от притока воды и ее расхода на испарение: чем больше приток, тем с большей площади зеркала вод она переходит в атмосферу под действием солнечного обогрева. Устойчивость Мирового океана обусловлена его огромной емкостью, что предупреждает значительные колебания объема океанических вод.
Продуктивность растительного покрова вначале обеспечивает ее медленный прирост, затем ускоряется и, при большой плотности, вновь замедляется из-за снижения интенсивности фотосинтеза в стесненных условиях. Соответственно, уменьшается и биомасса.
Растительноядные животные при большой численности испытывают затруднения с питанием, в связи с чем снижается коэффициент их рождаемости. Большая масса хищников начинает уничтожать не только больных и старых животных, содействуя повышению рождаемости растительноядных животных, но, превысив определенный предел, подрывает свою продуктивность (из-за уменьшения ресурсов питания).
Таким образом, экологические системы обладают определенной степенью устойчивости по отношению к тем отклонениям величин биомассы любого компонента системы от стационарного их состояния. При любых изменениях биомасс, превосходящих эти пределы, восстановление стационарных состояний полностью исключено, и один или несколько компонентов системы уничтожаются.
Устойчивость экологических систем, которые содержат определенные компоненты, обычно выше устойчивости систем, в которых некоторые группы животных отсутствуют.
Так как трофические взаимоотношения компонентов экосистем придают им определенную устойчивость, они обладают и определенными чертами целостности. Поэтому экологические системы могут оказывать влияние на процесс эволюции организмов.
Численность каждой группы животных, входящих в данную экосистему, постоянно колеблется. Она во многом определяется пищевыми ресурсами данной местности. Поэтому уничтожение животными источников собственного питания ведет к разрушению данной экосистемы и ставит под угрозу существование входящих в нее видов. Оставшиеся выжившими после разрушения прежней экологической системы группы организмов могут образовать новую экосистему с организмами из других систем.
В природных условиях коэффициенты рождаемости и смертности относительно неустойчивы. При определенном несоответствии их популяциям грозит вымирание. Поэтому успех живых организмов в борьбе за существование во многом определяется их способностью поддерживать устойчивость экосистем, в которые они входят.
Одновременно, само эволюционное развитие предполагает, что условия равновесия изменяются с течением времени: иначе была бы просто стабилизация существующего уровня организации экологических систем.
Следовательно, если даже наиболее устойчивые экосистемы эволюционируют, то это может происходить как под действием каких-то чисто биологических факторов, так и в результате изменений природной среды. Если эти изменения были более крупными - могло наступить вымирание многих групп живых организмов, с высвобождением определенных экологических ниш. Менее значимые нарушения устойчивости экосистем приводили также к вымиранию, но экологические ниши при этом сразу же заполнялись другими организмами. В этом отношении скорость эволюции должна каким-то образом коррелироваться с темпами изменений условий внешней среды. И между ними должно существовать определенное равновесие.
Именно высокая устойчивость экосистем свидетельствует, что крупномасштабные экологические катастрофы были редким явлением в геологической истории.
Таким образом, устойчивость биосферы определяется и определяет устойчивость экосистем, также как устойчивость последних определяется и определяет устойчивость популяций и т.д.
Как известно, в составе экологических дисциплин выделяюся аут- и синэкология. Первая из них изучает взаимоотношения между отдельными организмами, вторая - взаимоотношения организмов с окружающей средой. Поэтому синэкология во многом является и биогеографической наукой.
Основной биогеографический закон гласит: пространственная неоднородность географической оболочки отражается на пространственной неоднородности биосферы. Системе геохор В.Б. Сочавы соответствует адекватная ей система биохор. Геотопологический подход дает четкие критерии при оценки угодий под разные виды землепользования.
Постулатом ландшафтной геоботаники является утверждение, что один фитоценоз соответствует одному биогеоценозу, а последний одной фации.
Наиболее существенные свойства фитоценозов выступают в качестве индикаторов состояния данной среды. Они раскрываются через регистрацию видового состава и ценоэлементов растительных сообществ (рис. 7.3).
Соответственно, выделяются следующие фитоценотипы: эдификаторы, как строители главного яруса растительности; доминанты - виды преобладающие во второстепенных ярусах; второстепенные - редко встречающиеся, незначительные по массе растения.
| ЦЕНОЭЛЕМЕНТЫ | ||
| СОСТАВ | ||
Виды Синморфологические Синэкологические
Единицы единицы
Дата добавления: 2021-11-16; просмотров: 532;
Поиск по сайту
Узнать еще
- АНАЛИЗ ЧИСЛЕННОСТИ УПРАВЛЕНЧЕСКОГО ПЕРСОНАЛА ПО ПОДСИСТЕМАМ СИСТЕМЫ УПРАВЛЕНИЯ ОРГАНИЗАЦИИ
- АФИНСКАЯ МОРСКАЯ ДЕРЖАВА
- Биосферы и природных зон
- Вода как элемент биосферы. Значение воды для жизнеобеспечения человека.
- Воздействие на человека загрязнений биосферы
- Возникновение биосферы
- Г. Подсистема навигационной аппаратуры потребителей
- ГЕОХИМИЯ БИОСФЕРЫ. Структура биосферы

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
