Особенности строения и функционирования живых клеток
Как уже говорилось, биотехнология – это технология работы с биообъектами.
А основой строения любой живой материи, известной нам к настоящему времени, является клетка,
находящаяся в жизнедеятельном состоянии (исключая вирусы, более детально рассматривать которые
мы будем несколько позже – но которые также неспособны к размножению и вообще какой-либо
жизнедеятельности вне клетки–хозяина). Поэтому и изучение биотехнологии следует начинать
с рассмотрения особенностей строения и функционирования живых клеток.
Прежде всего любая такая клетка отделена от окружающей среды внешней цитоплазматической мембраной –
весьма избирательно проницаемой не только для высокомолекулярных химических веществ,
но даже для таких малых по размеру частиц, как ионы натрия или калия.
Вследствие чего химический состав внутренней среды живой клетки (называемой цитоплазмой), как правило, весьма значительно отличается от состава внешней среды, в которой находится рассматриваемая клетка –
являясь, к тому же, величиной не постоянной, а всё время и достаточно значительно меняющейся, вследствие процессов метаболизма, протекающих в клетке, характер которых также может в значительной степени меняться, в зависимости от стадии клеточного жизненного цикла, изменения условий внешней среды (включая не только её химический состав, но и температуру, освещение, давление и т.п.), а также вследствие разных иных причин (таких, например, как химические, механические, электромагнитные и иные сигналы от других клеток или вирусов).
При этом, цитоплазма представляет собой чрезвычайно многокомпонентный, жидкий, водный, коллоидный раствор, называемый ещё гиалоплазмой; включающий в себя большое количество различных биополимеров,
а также иных органических и неорганических веществ; и окружающий различные надмолекулярные клеточные органеллы и включения.
В свою очередь, цитоплазматическая мембрана (ЦПМ), по современным представлениям, состоит из достаточно регулярного двойного слоя липидов, способных ко взаимодвижению, но в основном, обращенных друг к другу своими гидрофобными частями (включающим в себя остатки высокомолекулярных жирных кислот), а внутрь и вне клетки – гидрофильными фрагментами (представляющими собой, как правило, глицериновый фрагмент). Плюс, в этот липидный бислой уже значительно менее регулярно включены различные белки (как интегральные, пронизывающие ЦПМ насквозь, так и периферические), а также олигосахариды. И таких белков и олигосахаридов главная роль которых состоит в избирательной сорбции (рецепторы) либо пропускании внутрь живой клетки и выводе из оной (ионные и другие транспортные каналы) различных химических веществ и их надмолекулярных агрегатов.
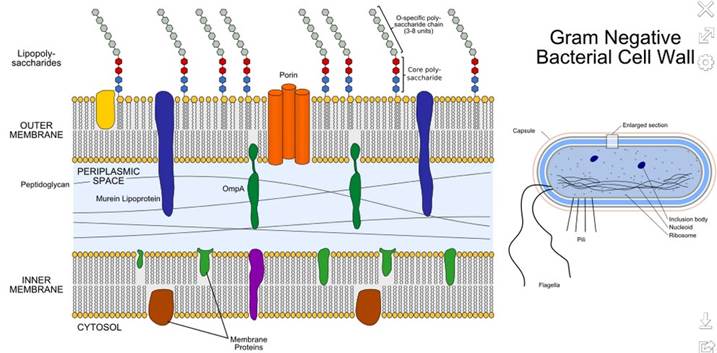
Второй важнейшей клеточной органеллой является клеточная стенка – представляющая собой расположенную снаружи от ЦПМ жёсткую оболочку клетки, наличествующую у большинства бактерий, грибов и растений, состоящую из различных углеводов, гликопротеинов и липополисахаридов (таких например, как муреин у бактерий, хитин у грибов, целлюлоза у растений и т.п.) и выполняющую различные дополнительные структурные, защитные и транспортные функции.
При этом если, к примеру, у грамположительных бактерий клеточная стенка представляет собой толстый слой пептидогликана, непосредственно покрывающий снаружи ЦПМ. То у грамотрицательных бактерий клеточная стенка представляет собой однослойный муреиновый мешок, поверх которого располагается ещё одна мембрана, сходная с цитоплазматической. Таким образом, у грамотрицательных бактерий между внутренней (плазматической) и наружной мембранами существует обособленный клеточный фрагмент (компартмент), называемый периплазматическим пространством (или периплазмой), в котором локализован целый ряд дополнительных ферментов, транспортных систем и т.п.
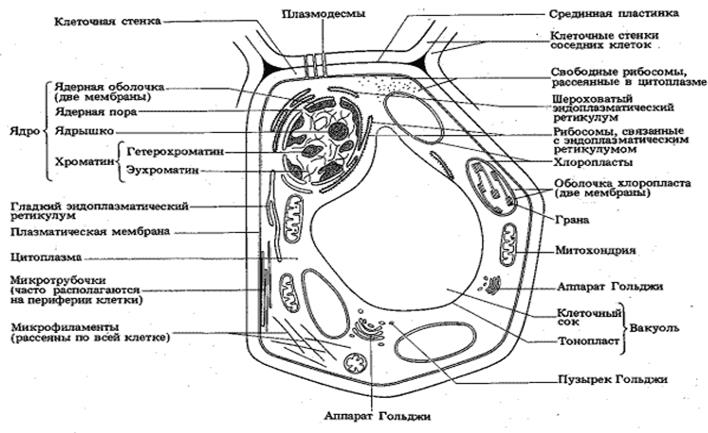
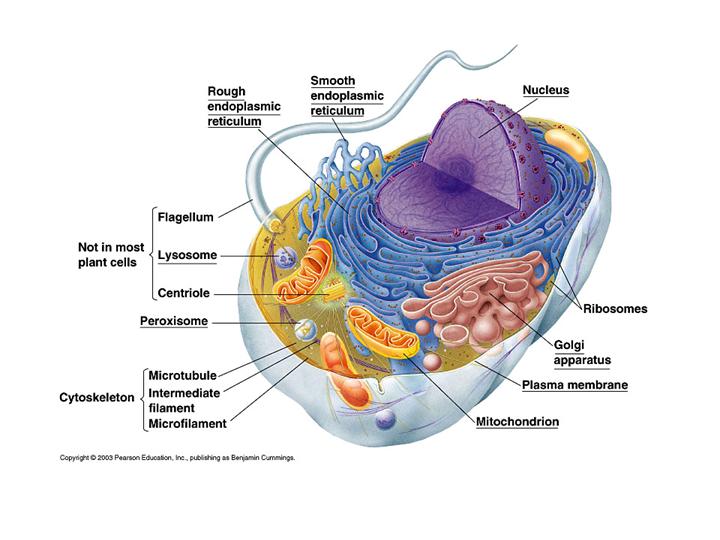
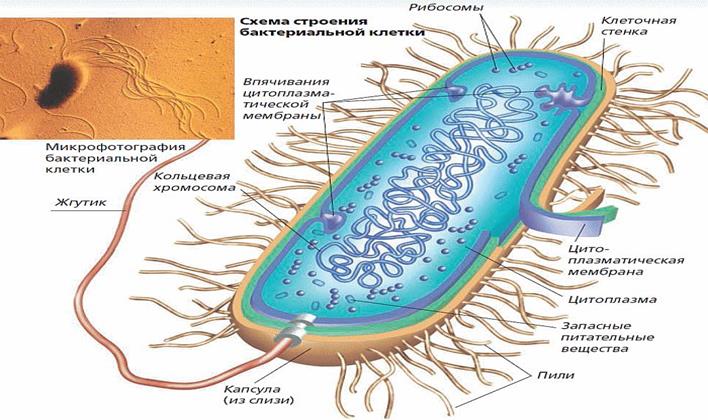
По наличию же других клеточных органелл все живые организмы делятся на прокариотов (к коим относятся бактерии – внутри клеток которых нет органелл, отделенных от основной гиалоплазмы собственной, дополнительной, избирательно проницаемой мембраной) и эукариотов (к коим относятся простейшие, грибы, растения и животные).
При этом, из основных внутриклеточных органелл следует упомянуть, прежде всего:
(1) ядро – выполняющее у эукариотов функции хранения и воспроизведения генетической информации
и окруженное собственными двумя мембранами (внешняя из которых через эндоплазматическую сеть связана с ЦПМ);
(2) затем митохондрии – являющиеся энергетическими центрами эукариотических клеток, обеспечивающими генерацию электрического потенциала и синтез молекул АТФ (аденозин трифосфата) в результате процессов многостадийного окислительного фосфорилирования, называемых «клеточным дыханием», в ходе которого молекулы кислорода преобразуются в углекислый газ»; а энергия, выделяющаяся при расщеплении молекул АТФ, в дальнейшем, может использоваться клеткой для осуществления большинства биохимических реакций; кроме того, внутри митохондрий локализуется ряд других важных для клетки биохимических реакций; а сами митохондрии, также как и клеточное ядро, окружены двумя мембранами (причем, внутренняя мембрана по своему составу отличается от внешней), плюс имеют собственную ДНК (отличающуюся от ядерной) – так что по современным представлениям митохондрии исходно являлись самостоятельными прокариотическими микроорганизмами, поглощенными эукариотической клеткой–хозяйкой и наладившими затем с ней симбиотические отношения (вместо того, чтобы быть переваренными ею, как в случае например амёб, охотящихся на бактерий);
(3)эндоплазматическую сеть (иначе, ретикулум или ЭПР) – представляющую собой разветвлённую систему окружённых мембраной (морфологически идентичной внешней оболочке клеточного ядра и составляющей с ней одно целое) уплощённых полостей, на поверхности которых, в частности, локализуются рибосомы (представляющие собой сложные нуклеопротеидные комплексы, с помощью которых внутри живой клетки осуществляется синтез различных белков) – вследствие чего ЭПР играет важную роль в синтезе, модификации (приводящей, в частности, к нейтрализации многих опасных для клетки токсинов) и транспорте внутриклеточных белков, липидов, полисахаридов и ряда других биополимеров;
(4)комплекс Гольджи – представляющий собой стопку дискообразных мембранных мешочков, предназначенных для выведения из клетки и распределения внутри неё веществ, синтезированных на поверхности ЭПР, а также образования новых внутриклеточных мембран;
(5)лизосомы – представляющие собой отдельные мембранные пузырьки, ответственные за разрушение и трансформацию внутри клетки различных биополимеров, как синтезированных ранее самой клеткой, так и поступивших в неё из вне;
(6)вакуоли – аналоги лизосом, служащие для секреции, экскреции и хранения внутри клетки различных веществ;
(7) различные виды протопластов (иначе, пластид) – представляющие собой аналоги митохондрий (тоже окруженные двумя мембранами, имеющие собственную ДНК и т.д.), но выполняющие внутри растительной эукариотической клетки–хозяйки другие функции – такие как хлоропласты (содержащие хлорофилл, с помощью которого растительные клетки осуществляют процессы фотосинтеза, в результате коих из молекул воды и углекислого газа с помощью световой энергии происходит образование различных высокомолекулярных биоорганических соединений), хромопласты, лейкопласты и т.п.;
(8) и наконец, жгутики, пили и т.п. – обеспечивающие подвижность одноклеточных микроорганизмов и некоторых видов клеток в многоклеточных организмах.

Жизнь – принципиально неравновесный процесс. Поэтому любой организм (в т.ч. одноклеточный), находящийся в жизнедеятельном состоянии, постоянно обменивается с окружающей средой множеством различных химических веществ, которые внутри него ещё и многократно преобразуются.
При этом, первой стадией таких процессов является транспорт того или иного вещества внутрь клетки. Причем, если прокариотические микроорганизмы, грибы и водоросли могут транспортировать внутрь клетки только вещества, растворенные в воде. То некоторые эукариоты способны поглощать из окружающей среды нерастворимые вещества в виде твердых кусочков или капель жидкостей, не смешивающихся с водой. Этот процесс носит название эндоцитоза и подразделяется на фагоцитоз (при котором поглощаются твердые вещества) и пиноцитоз (при котором поглощаются жидкости).
При эндоцитозе участок ЦПМ рядом с частицей пищи втягивается внутрь и замыкается, образуя вокруг вышеупомянутой частицы вакуоль. Эта вакуоль, находясь в цитоплазме, сливается с лизосомой, содержащей набор гидролитических ферментов. В образовавшейся пищеварительной вакуоли частица пищи лизируется. Растворимые продукты гидролиза диффундируют в цитоплазму, а неразрушенные остатки экскретируются наружу путем, обратным эндоцитозу. Среди микроорганизмов эндоцитоз могут осуществлять некоторые простейшие. А у высших животных эндоцитоз сохранился в виде защитной функции поглощения чужеродных клеток фагоцитами, а обратный эндоцитозу процесс – при выделении гормонов.
Также, поскольку у бактерий, грибов и растений клеточная стенка является существенным препятствием для высокомолекулярных веществ – то перед транспортом таких веществ внутрь клетки они сначала расщепляются на олиго- и мономеры вне оной клетки соответствующими ферментами (экзогидролазами), выделяемыми клеткой либо непосредственно во внешнюю среду, либо на наружную сторону её ЦПМ.
Кроме того, у грамотрицательных бактерий экзогидролазы могут быть локализованы в периплазматическом пространстве. И высокомолекулярные вещества в этом случае проникают в периплазму через поры, образованные белками–поринами, находящимися в наружной мембране.
В целом, у живых организмов существует несколько принципиально различных способов поступления веществ внутрь клетки. Так, во-первых, все незаряженные молекулы (включая воду, газы и т.п.) могут поступать в клетку путем пассивной диффузии. В этом случае вещества проникают в клетку по градиенту концентрации. Данный процесс не требует затрат энергии. Но происходит, как правило, с весьма невысокой скоростью и лишь до тех пор, пока не наступит равновесие между содержанием вещества вне и внутри клетки.
При облегченной диффузии скорость транспорта веществ внутрь клетки значительно увеличивается за счёт участия в этом процессе специфических белков–переносчиков, называемых пермеазами. При этом, пермеазы либо расположены поперек мембраны в виде интегральных белков, либо способны передвигаться через мембрану как в свободном, так и в связанном с транспортируемым веществом состоянии. Облегченная диффузия также не требует энергетических затрат и продолжается, пока есть градиент концентраций транспортируемых веществ. Эукариотические клетки путем облегченной диффузии транспортируют различные сахара и аминокислоты. А у микроорганизмов путем облегченной диффузии происходит также, как правило, экскреция, т.е. выделение наружу из клетки различных продуктов её метаболизма.
Поступление же веществ в клетку (или обратный этому процесс экскреции) против градиента концентрации может происходить только благодаря механизмам активного транспорта. Такие механизмы требуют затрат энергии, запасаемой клеткой в виде молекул АТФ, и специальных весьма сложных белковых транспортных систем.
Так, в частности, при работе многих мембранных ферментов происходит выброс протонов (Н+ ионов) во внешнюю среду, в результате чего образуется трансмембранный градиент протонов. Этот процесс носит название первичного транспорта. Протонный градиент в клетке всегда поддерживается на определенном уровне с помощью дыхания, фотосинтеза, брожения. Активный же транспорт называют вторичным – поскольку он происходит за счет вышеупомянутого трансмембранного градиента протонов. И в целом, известно несколько видов такого активного транспорта. При симпорте в одном направлении переносятся одновременно два вещества, при унипорте – одно, а антипорт предполагает движение двух веществ в противоположных направлениях. Кроме того, грамотрицательные бактерии из-за наличия наружной мембраны имеют дополнительные компоненты транспортных систем для переноса вещества через своё периплазматическое пространство. Эти высокоспецифичные растворимые белки образуют комплекс с субстратом на внутренней стороне внешней мембраны и пересекают периплазму, доставляя его на соответствующие пермеазы, которые с затратой энергии транспортируют субстрат внутрь клетки. Транспортные системы с участием связывающих белков имеются и у некоторых грамположительных микроорганизмов, но тогда связывающие белки «заякорены» своей N-концевой частью в ЦПМ.
Далее, процесс преобразования поступающих из вне химических веществ осуществляется уже внутри клетки. Этот процесс называется метаболизмом и подразделяется на катаболизм (приводящий к разложению различных веществ на более простые составляющие, сопровождаемому, как правило, выделением энергии – которая, также как и при «клеточном дыхании» может запасаться клетками в виде молекул АТФ) и анаболизм (приводящий к синтезу различных необходимых клетке веществ из более простых составляющих, требующему, как правило, затрат энергии, получаемой клеткой за счет отщепления от молекулы АТФ одной или 2-х фосфатных групп).
При этом, основную роль в процессах метаболизма играют белки, выполняющие функцию биокатализаторов, называемых также ферментами или энзимами, без которых невозможно осуществление практически ни одной из биохимических реакций, важных как для жизнедеятельности самих биообъектов, используемых в биотехнологии, так и для получения целевых продуктов в любых биотехнологических процессах.
Причем, за счет действия этих ферментов биохимические реакции по сравнению с обычными химическими, как уже говорилось, могут протекать:
(1) со значительно более высоким выходом по целевым продуктам
+ (2) значительно более избирательно и специфично
+ (3) а также в существенно более мягких условиях (в т.ч. в плане энергозатрат).
Вследствие чего, многие биохимические реакции обычным химическим путем провести просто невозможно
либо весьма трудно.
Дата добавления: 2017-10-04; просмотров: 2075;
Поиск по сайту
Узнать еще
- D-технология построения чертежа. Типовые объемные тела: призма, цилиндр, конус, сфера, тор, клин. Построение тел выдавливанием и вращением. Разрезы, сечения.
- F3 Аффективные расстройства настроения
- F38 Другие (аффективные) расстройства настроения
- I. ОСОБЕННОСТИ ДЕЛОВОГО И ЛИЧНОСТНОГО ОБЩЕНИЯ В СОВМЕСТНОЙ ДЕЯТЕЛЬНОСТИ
- I.2. Основные категории водопотребления промышленных предприятий и их особенности
- I2. Особенности аэродинамики несущего винта (НВ)
- II. Завоевание Китая маньчжурами. Экономическое положение страны в XVII – начале XIX вв.: аграрная политика Цинской династии, особенности развития городского ремесла
- II. Особенности политического устройства Ирана

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
