Примеры орудийных действий животных разных видов в природе и в экспериментах
| Инстинкты | «Инсайт» | Обучение и традиции | |
| Дятловые вьюрки: | Шимпанзе: | Шимпанзе: | |
| добывание насекомых с помощью палочек | — сооружение вышек; | — «ужение» термитов; | |
| — угроза канистрами; | |||
| — применение палок; | — разбивание орехов камнями на наковальнях; | ||
| Калифорнийские каланы: | — побеги из клеток; | ||
| — тушение огня | |||
| разбивание раковин камнями | Сойки: | Макаки: | |
| изготовление бумажных «жгутов» для доставания пищи | — мойщики картофеля ; | ||
| Новокаледонские галки: | |||
| — крабоеды | |||
| изготовление «крючков» для ловли насекомых | |||
В табл. 4.1 приведены некоторые примеры орудийных действий животных, имеющих разную природу. Они иллюстрируют представление о том, что в основе сходных по внешнему проявлению поведенческих актов могут лежать разные механизмы — инстинкт, обучение, разум (см. 1.2). Удельный вес каждого из этих факторов может быть разным в зависимости от многих обстоятельств, даже если конечный результат внешне выглядит одинаково (Крушинский, 1986). Поэтому (в соответствии с «каноном К. Л. Моргана») то или иное проявление орудийной деятельности может быть отнесено кактам мышления только после тщательного анализа.
Эти факты демонстрируют сложность и неоднородность такой формы поведения животных, как орудийная деятельность. В основе этих актов, схожих по внешнему проявлению, могут лежать принципиально разные механизмы:
♦ экстренное решение в новой ситуации («инсайт»);
♦ обучение методом проб и ошибок и подражание сородичам;
♦ выполнение видоспецифической (инстинктивной) программы.
4.6. Методики, разработанные Л. В. Крушинским для изучения способности животных к поиску приманки, исчезающей из поля зрения.
Рассмотренные выше эксперименты убедительно показали способность антропоидов к целенаправленному употреблению орудий в соответствии с «мысленным планом». Данную способность можно рассматривать как проявление элементарного мышления.
В то же время описанные выше методы анализа решения животными задач имели определенные ограничения:
♦ результаты таких опытов носили чисто описательный характер, и субъективизм в их трактовке был почти неизбежен;
♦ при повторении эксперимента неизменно возникал вопрос о том, что животное не решает задачу заново, а стереотипно применяет опыт, приобретенный в предшествующей пробе;
♦ такие методики практически невозможно было использовать в опытах на животных-неприматах, а поэтому исключалась возможность сравнительного анализа, необходимого для ответа на вопрос, насколько широко зачатки мышления представлены у более примитивно организованных животных (см. 1.3).
Ответ на последний вопрос требовал другого методологического подхода. Для его изучения нужны были универсальные тесты, которые можно было бы предлагать разным животным и при этом получать результаты, пригодные для количественной оценки, статистической обработки и получения сравнительной характеристики разных видов.
Такие методологические подходы были созданы независимо друг от друга двумя учеными — Г. Харлоу и Л. В. Крушинским. Г. Харлоу (см. 3.3.2) в 50-е годы предложил метод сравнительной оценки высших когнитивных функций животных, который дает возможность выяснить, улавливают ли животные общий принцип, лежащий в основе их решения, т.е., по выражению автора, формируется ли у них «установка на обучение».
Попытка Г. Харлоу была удачной. С помощью его теста действительно можно было почти в стандартных условиях исследовать самых разных животных и охарактеризовать динамику их обучения количественными параметрами. Однако метод Харлоу позволял охарактеризовать в основном одну сторону мышления животных — способность к обобщению.
Л. В. Крушинский предложил универсальные методики тестирования и предпринял широкое сравнительное исследование способности животных к другому виду элементарного мышления — решению экстренно возникших задач, для которых у них нет готовой программы.
В отличие от описанных выше задач, в которых надо было достать удаленную, но видимую цель, значительная часть методик, предложенных Л. В. Крушинским для изучения зачатков мышления животных, основана на поиске приманки, тем или иным способом исчезающей из поля зрения, как только животное начинало ее есть. Этим они отличались от ранее рассмотренных методик, в которых цель всегда была «в пределах зрительного поля». Поэтому решение задач в методиках Кру-шинского должно было осуществляться не под контролем внешних стимулов, а за счет оперирования «образом исчезнувшей приманки» (Более подробно методики описаны в книге Л. В. Крушинского «Биологические основы рассудочной деятельности» (1986)..
Повторим определение мышления (рассудочной деятельности животных), данное Л. В. Крушинским, которое он называл «рабочим»:
«Способность животного улавливать эмпирические законы, связывающие предметы и явления внешнего мира, и оперировать этими законами в новой для него ситуации для построения программы адаптивного поведенческого акта».
4.6.1. Понятие об «эмпирических законах» и элементарной логической задаче.
Л. В. Крушинский ввел понятие элементарной логической задачи, т.е. задачи, которая характеризуется смысловой связью между составляющими ее элементами. Благодаря этому она может быть решена экстренно, при первом же предъявлении, за счет мысленного анализа ее условий. Такие задачи по своей природе не требуют предварительных проб с неизбежными ошибками. Как и задачи, которые рассматривались в разд. 4.5, они могут служить альтернативой и «проблемному ящику» Торндайка, и выработке различных систем дифференцировочных УР. Дифференцировочные УР, точнее, порядок смены подкрепления по ходу их выработки, не связаны для животного с «естественной» логикой событий внешнего мира. Именно поэтому их нельзя решить иначе, чем постепенно, при многократных пробах и подкреплении правильных реакций.
Отличительную черту тестов Крушинского составляет то, что их решение требует (согласно его определению) оперирования так называемыми «эмпирическими законами». Имеется в виду, что анализу подлежат такие свойства предметов и явлений, которые присущи им в силу природных физических законов и с которыми животное постоянно сталкивается в жизни.
Прибегая к терминологии когнитивной психологии, можно сказать, что эти «законы» входят в состав «познавательной карты», или «образной картины мира животного», т.е. той системы знаний, которую оно накапливает в течение жизни. На необходимость и плодотворность использования таких тестов указывал Д. Примэк (Premack, 1983), называя эту форму мышления животных «естественным мышлением» (natural reasoning), в отличие от их способности к обобщению и умозаключениям.
Тесты для изучения способности к обобщению и умозаключению организованы таким образом, что их условия и структура достаточно произвольно определяются экспериментатором и совершенно не связаны с естественными закономерностями. Это относится также к тесту Ревеша-Крушинского, где алгоритм изменений положения приманки задается экспериментатором произвольно и не имеет никакой связи с процессами в естественной для животных среде (см. 4.7), или формированию установки на обучение в тестах Харлоу.
Базовые формы когнитивной деятельности животных были частично описаны в гл. 3. Ниже перечислены наиболее важные эмпирические законы, владение которыми, как писал Л. В. Крущинский, необходимо животному для решения ряда логических задач.
1. Закон «неисчезаемости» предметов (object permanence). Животные способны сохранять память о предмете, ставшем недоступным непосредственному восприятию. Животные, «знающие» этот эмпирический закон, более или менее настойчиво ищут корм, тем или иным способом скрывшийся из их поля зрения (описание поведения млекопитающих в тесте на отсроченную реакцию см. в 3.4.1). Представление о «неисчезаемости» есть и у многих птиц. Так, вороны и попугаи активно ищут корм, который у них на глазах накрыли непрозрачным стаканом или отгородили от них непрозрачной преградой. В отличие от этих птиц голуби и куры законом «неисчезаемости» не оперируют или оперируют в весьма ограниченной степени. Это выражается в том, что в большинстве случаев они почти не пытаются искать корм после того, как перестают его видеть.
Представление о «неисчезаемости» предметов необходимо для решения всех типов задач, связанных с поиском приманки, скрывшейся из поля зрения.
2. Закон, связанный с движением, — одним из самых универсальных явлений окружающего мира, с которым сталкивается любое животное независимо от образа жизни. Каждое из них без исключения с первых же дней жизни наблюдает перемещения родителей и сибсов, хищников, которые им угрожают, или, наоборот, собственных жертв.
Вместе с тем животные воспринимают изменения положения деревьев, травы и окружающих предметов при собственных перемещениях.
Это создает основу для формирования представления о том, что движение предмета всегда имеет определенное направление и траекторию.
Знание этого закона лежит в основе решения задачи на экстраполяцию (см. 4.5.5.2).
3. Законы «вмещаемости» и «перемещаемости». Имеется в виду, что на основе восприятия и анализа пространственно-геометрических признаков окружающих предметов животные «знают», что одни объемные предметы могут вмещать в себя другие объемные предметы и перемещаться вместе с ними.
В лаборатории Л. В. Крушинского разработаны две группы тестов, с помощью которых можно оценивать способность животных разных видов оперировать указанными эмпирическими законами в новой ситуации. Схемы этих экспериментов представлены ниже (см. 4.6.3).
Как полагал Крушинский, перечисленные имзаконы не исчерпывают всего, что может быть доступно животным. Он допускал, что они оперируют также представлениями о временных и количественных параметрах среды, и планировал создание соответствующих тестов. Животные действительно способны оценивать количественные и даже числовые параметры стимулов (см. гл. 5).
Многие животные, в том числе обезьяны, в тесте на предпочтение выбирают стимулы большей площади и объема. По-видимому, они могут, также без специальной тренировки, воспринимать и сравнивать стимулы, различающиеся по числу элементов. Вороны и голуби, например, без всякой предварительной подготовки выбирают кормушку, содержащую большее число зерен или личинок мучного хрущака. В обоих случаях тест проводится в ситуации «свободного выбора», когда птицы съедают любую выбранную ими приманку, а обезьяны получают любой выбранный ими стимул.
Предложенные Л. В. Крушинским (1986) и описанные ниже методики сравнительного изучения рассудочной деятельности с помощью элементарных логических задачоснованы на допущении, что животные улавливают эти «законы» и могут использовать их в новой ситуации. Задачи построены так, что возможно их экстренное решение, принятое логическим путем, в соответствии с лежащим в их основе принципом, и не требующее предварительного обучения по методу проб и ошибок.
4.6.2. Методика изучения способности животных к экстраполяции направления движения пищевого раздражителя, исчезающего из поля зрения («задача на экстраполяцию»).
Под экстраполяцией понимают способность животного выносить функцию, известную на отрезке, за ее пределы. Л. В. Крушинский предложил несколько элементарных логических задач для изучения такой способности.
Наибольшее распространение получил так называемый «опыт с ширмой» (рис. 4.12). В этом опыте перед животным помещают непрозрачную преграду — ширму (длина — около 3 м, высота 1 м). В центре ширмы имеется вертикальная щель, через которую видны две кормушки, в начале опыта расположенные прямо перед щелью. Кормушки разъезжаются в стороны, как только животное начинает есть, но оно может видеть начальный участок их пути до момента исчезновения за поперечными преградами-клапанами. Через несколько секунд кормушки скрываются из поля зрения, так что их дальнейшее перемещение животное уже не видит и может только представлять его мысленно.
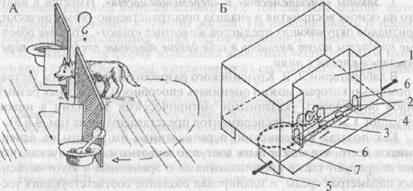
Рис. 4.12. Исследование способности к экстраполяции в опытах с ширмой.
А — общий вид установки для опытов с хищными млекопитающими, кроликами и птицами (рисунок Т. Никитиной); Б — камера для экспериментов с мышами (пояснение в тексте).
За щелью находятся две кормушки: одна с кормом, другая пустая. Это делается для того, чтобы создать животному возможность альтернативного выбора. К тому же если двигаются две кормушки, то животное не сможет отыскать корм, ориентируясь на звук при движении.
Опыты с грызунами проводятся несколько по-другому. Вторую кормушку тоже наполняют кормом (поилки с молоком). Эту кормушку либо закрывают сеткой (в опытах с крысами), либо (опыты с мышами) ставят так, чтобы животное ее не видело. Это позволяет «уравнять запахи», идущие от приманки с двух сторон камеры, и тем самым препятствовать отысканию корма с помощью обоняния. Экспериментальная камера для исследования способности к экстраполяции у мышей (рис. 4.12Б) устроена так, чтобы животное оставалось в ней в интерватах между предъявлениями задачи. В одной из стенок камеры (1) на середине ее длины на уровне пола имеется отверстие (2), дающее животному доступ к поилке (3) и позволяющее видеть начальный этап ее перемещения. Контрольная поилка (4) перемещается в противоположную сторону. С помощью специального рычага (5) их можно передвинуть к боковым отверстиям (6). «Обход ширмы» в этом варианте установки — это перемещение животного вправо или влево и подход к одному из боковых отверстий. Траектория (7) показывает путь мыши при правильном решении задачи.
Чтобы решить задачу на экстраполяцию, животное должно представить себе траектории движения обеих кормушек после исчезновения из поля зрения и на основе их сопоставления определить, с какой стороны надо обойти ширму, чтобы получить корм.
Способность к решению этой задачи проявляется у многих позвоночных, но ее выраженность значительно варьирует у разных видов.
Основной характеристикой способности животных к рассудочной деятельности служат результаты первого предъявления задачи, потому что при их повторении подключается влияние и некоторых других факторов. В связи с этим для оценки способности к решению логической задачи у животных данного вида необходимо и достаточно провести по одному опыту на большой группе. Если доля особей, правильно решивших задачу при ее первом предъявлении, достоверно превышает случайный уровень, считается, что у животных данного вида или генетической группы есть способность к экстраполяции (или к другому виду рассудочной деятельности).
Как показали исследования Л. В. Крушинского, животные многих видов (хищные млекопитающие, дельфины, врановые птицы, черепахи, крысы-пасюки, мыши некоторых генетических групп) успешно решали задачу на экстраполяцию. В то же время животные других видов (рыбы, амфибии, куры, голуби, большинство грызунов) обходили ширму чисто случайно (рис. 4.13А).
При повторных предъявлениях задачи поведение животного зависит не только от способности (или неспособности) экстраполировать направление движения, но и от того, запомнило ли оно результаты предыдущих решений. Ввиду этого данные повторных опытов отражают взаимодействие ряда факторов, и для характеристики способности животных данной группы к экстраполяции их надо учитывать с известными оговорками.
Многократные предъявления позволяют точнее проанализировать поведение в опыте животных тех видов, которые плохо решают задачу на экстраполяцию при ее первом предъявлении (о чем можно судить по невысокой доле правильных решений, которая не отличается от случайного 50%-го уровня). Оказывается, что большинство таких особей ведет себя чисто случайным образом и при повторениях задачи. При очень большом числе предъявлений (до 150) такие животные, как, например, куры или лабораторные крысы, постепенно обучаются чаще обходить ширму с той стороны, в которую скрылся корм. Напротив, у хорошо экстраполирующих видов результаты повторных применений задачи могут быть несколько ниже, чем результаты первого, хотя продолжают достоверно превышать случайный уровень (например, у лисиц и собак, рис. 4.13Б). Причиной такого снижения показателей теста может быть, по-видимому, влияние различных тенденций в поведении, напрямую не связанных со способностью к экстраполяции как таковой. К ним относится склонность к спонтанному чередованию побежек, предпочтение одной из сторон установки, характерное для многих животных, и т.д. В опытах Крушинского и сотрудников у некоторых животных (например, врановых и хищных млекопитающих) обнаруживались проявления боязни обстановки опыта, нередко сопровождавшиеся беспорядочными обходами ширмы независимо от направления движения корма.
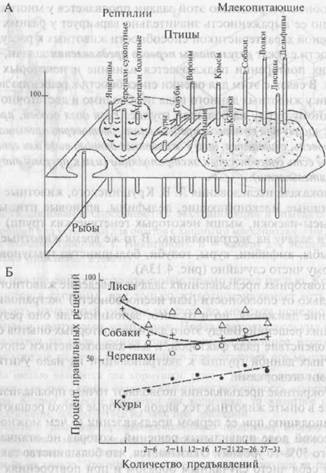
Рис. 4.13.Решение задачи на экстраполяцию.
А — успешность решения задачи на экстраполяцию животными разных таксономических групп при ее первом предъявлении; по оси ординат — доля правильных решений в процентах (плоскость соответствует 50%-му случайному уровню правильных решений); Б — усредненные кривые успеха решения задачи при ее многократных предъявлениях животными разных таксономических групп (по Крушинскому, 1986). По оси ординат — как в случае А, по оси абсцисс — номера предъявлений.
Вопрос о влиянии разных стратегий поведения в опыте на проявление способности к экстраполяции был подробно проанализирован на уникальной модели — двух парах линий мышей с робертсоновской транслокацией на разном генетическом фоне (Полетаева, 1998). Оказалось, что эта способность зависит иот генотипа (см. также гл. 9).
С помощью теста на экстраполяцию, который позволяет давать точную количественную оценку результатов его решения, впервые была дана широкая сравнительная характеристика развития зачатков мышления у позвоночных всех основных таксономических групп, изучены их морфофизиологические основы, некоторые аспекты формирования в процессе онто- и филогенеза, т.е. практически весь тот круг вопросов, ответ на которые, согласно Н. Тинбергену (Tinbergen, 1963), необходим для всестороннего описания поведения.
4.6.3. Методики изучения способности животных к оперированию пространственно-геометрическими признаками предметов.
Анализ пространственных характеристик необходим во многих ситуациях, с которыми сталкиваются животные в естественной среде обитания. В опытах Толмена (1997) была продемонстрирована способность животных к обучению в лабиринте за счет формирования и запоминания мысленной «пространственной карты» (см. 3.4). Эту способность интенсивно изучают в настоящее время.
Элементы пространственного мышления обезьян были обнаружены и в опытах В. Келера. Он отмечал, что во многих случаях, намечая путь достижения приманки, обезьяны предварительно сопоставляли, как бы «оценивали» расстояние до нее и высоту предлагаемых для «строительства» ящиков. Понимание пространственных соотношений между предметами и их частями составляет необходимый элемент более сложных форм орудийной и конструктивной деятельности шимпанзе (Ладыгина-Котс, 1959; Фирсов, 1987).
К пространственным признакам можно отнести также геометрические свойства предметов (например, форму, наличие или отсутствие симметрии, размерность). С их анализом связаны эмпирические законы «вмещаемости» и «перемещаемости» объемных (трехмерных) предметов, которые могут вмещать друг друга и перемешаться, находясь один в другом.
Задача на оперирование эмпирической размерностью фигур (ОЭРФ).Л. В. Крушинский (1986) предложил тест для оценки одной из форм пространственного мышления — способности животного в поисках приманки сопоставлять предметы разной размерности: трехмерные (объемные) и двумерные (плоские).
Суть теста состоит в том, что объемная приманка может быть помещена (и спрятана) только в объемную (ОФ), но не в плоскую (ПФ) фигуру, поэтому животное должно выбрать ОФ.
Он был назван тестом на «оперирование эмпирической размерностью фигур» или тестом на «размерность» (Дашевский, 1977; 1979).
Этот термин был введен для характеристики предлагаемой задачи потому, что так называемая «плоская фигура», хотя и имела минимальную толщину, на самом деле также была трехмерной. Однако поскольку соотношение толщины плоской фигуры и размера «в глубину» объемной фигуры было от 1:40 до 1:100, то при предъявлении в паре такие фигуры имели четко различную «пространственность» и эмпирически оценивались как фигуры разной размерности. Форму фигур, которые давались животному для сопоставления, подбирали так, чтобы плоская была фронтальной проекцией объемной.
Для успешного решения задачи на ОЭРФ животные должны владеть следующими эмпирическими законами и выполнять следующие операции:
♦ мысленно представить себе, что приманка, ставшая недоступ
ной для непосредственного восприятия, не исчезает (закон «не-
исчезаемости»), а может быть помещена в другой объемный
предмет и вместе с ним перемещаться в пространстве (законы
«вмещаемости» и «перемещаемости»);
♦ оценить пространственные характеристики фигур;
♦ пользуясь образом исчезнувшей приманки как эталоном, мысленно сопоставить эти характеристики между собой и решить, где спрятана приманка;
♦ сбросить объемную фигуру и овладеть приманкой.
Первоначально опыты были проведены на собаках, но методика экспериментов была сложна и непригодна для сравнительных исследований. Б. А. Дашевский (1972) сконструировал установку, применимую для исследования этой способности у любых видов позвоночных, включая человека.
Она представляет собой стол, в средней части которого расположено устройство для раздвигания вращающихся демонстрационных платформ с фигурами. Животное находится по одну сторону стола, фигуры отделены от него прозрачной перегородкой с вертикальной щелью в середине. По другую сторону стола находится экспериментатор. В части опытов животные не видели экспериментатора: он был скрыт от них за перегородкой из стекла с односторонней видимостью.
Опыт ставится следующим образом (рис. 4.14). Голодному животному предлагают приманку (1), которую затем прячут за непрозрачный экран-коробку (2). Под его прикрытием приманку помещают в объемную фигуру (ОФ), например куб, а рядом помещают плоскую фигуру (ПФ), в данном случае квадрат (проекцию куба на плоскость). Затем экран удаляют, и обе фигуры, вращаясь вокруг собственной оси, раздвигаются в противоположные стороны с помощью специального устройства (3). Чтобы получить приманку, животное должно опрокинуть объемную фигуру (4).
Процедура эксперимента позволяла многократно предъявлять задачу одному и тому же животному, но при этом обеспечивать максимально возможную новизну каждого предъявления.
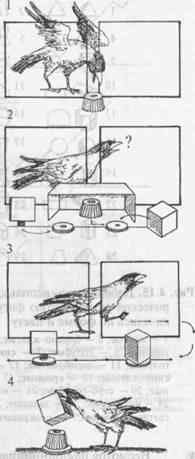
Рис. 4.14. Эксперимент с вороной по оперированию эмпирической размерностью фигур (рисунок Т. Никитиной).
Для этого всякий раз животному предлагали новую пару фигур, отличающуюся от остальных по цвету, форме, размеру, способу построения (плоскогранные и тела вращения) и размеру (рис. 4.15).
Примеры индивидуальных «кривых накопления», демонстрирующих динамику успешности решения задачи на ОЭРФ, даны на рис. 4.16. На этих кривых правильное решение задачи — выбор объемной фигуры — изображен отрезком прямой, направленным под углом 45° вверх по оси абсцисс, выбор плоской фигуры — таким же «шагом вниз», отсутствие выбора — горизонтальным отрезком. Обезьяны, дельфины, медведи и врановые птицы успешно решают эту задачу. Как при первом предъявлении теста, так и при повторных пробах они выбирают преимущественно объемную фигуру. В отличие от них хищные млекопитающие и часть врановых птиц реагируют на фигуры чисто случайно и лишь после десятков сочетаний постепенно обучаются правильным выборам.
Эти эксперименты позволили существенно уточнить картину различий в уровнях развития зачатков мышления у животных разных таксономических групп.
Особое значение имеет факт сходства в решении этого теста у врановых птиц и наиболее высокоорганизованных млекопитающих — низших узконосых обезьян, дельфинов, а также медведей, тогда как большинство других хищных млекопитающих его не решает. Такие же различия между ними были обнаружены по показателям формирования установки на обучение (см. 3.3.3) и довербальных понятий (см. 5.5.4).

Рис. 4.15. Набор фигур, использованных в тесте на оперирование эмпирической размерностью фигур (по Дашевскому, 1972). Фигуры различались по форме и цвету.
1 — желтые; 2 — бледно-желтые; 3 — темно-серые; 4 — зеленые; 5 — неокрашенные; 6 — голубые; 7 — синие; 8 — темно-зеленые; 9 — желтые; 10— голубые; 11 — серебристые; 12 — зеленые; 13— серые; 14 — бордовые; 15 — сине-зеленые; 16 — красные; 17 — оранжевые; 18 — светло-серые; 19 — черные; 20 — серо-голубые; 21 — малиновые; 22 — темно-розовые; 23 — белые; 24 — малиновые; 25 — золотые; 26 — фиолетовые; 27 — неокрашенные; 28 — светло-розовые; 29 — неокрашенные; 30 — черные.
Несмотря на принципиальные различия в строении мозга млекопитающих и птиц (отсутствие у птиц новой коры), наиболее высокоразвитые представители обоих классов достигают сходных, достаточно высоких уровней развития элементарного мышления.
Контрольный опыт.Схема задачи на оперирование размерностью позволила на ее основе разработать принципиально важный контрольный эксперимент — альтернативу логической задачи. При этом все «внешние атрибуты» опыта сохраняются, за исключением собственно логической структуры теста.
Задачу, предлагаемую вподобном контрольном эксперименте, нельзя решить при первом предъявлении за счет «понимания» ее смысла. То, какой выбор является правильным, можно установить только по ходу последовательных предъявлений теста (Дашевский, 1979). Поясним это на примере, приведенном на рис. 4.17. Слева вверху (А) (как и на рис. 4.14) дана схема исходного теста на ОЭРФ. В контрольном опыте (Б, В) демонстрационные платформы (2), на которых в собственно эксперименте на ОЭРФ животному показывали приманку (1), а затем устанавливали фигуры (4 и 5), заменены кормушками такого же диаметра (3). Подкрепление можно помещать в любую из кормушек, и ее можно накрыть крышкой с прикрепленной к ней ОФ (как на рис. 4.17Б) или ПФ (как на рис. 4.17В).
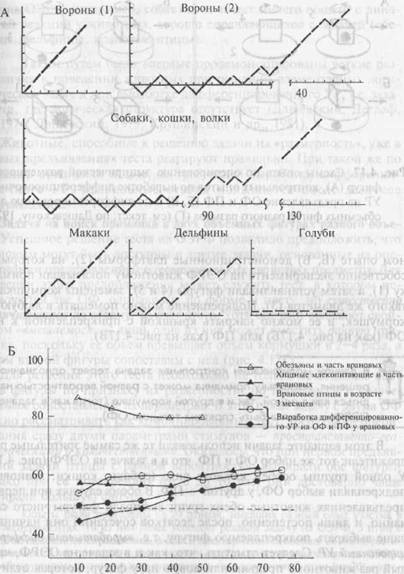
Рис. 4.16. Успешность решения задачи на оперирование эмпирической размерностью фигур животными разных видов.
А — примеры «кривых накопления». По оси ординат — разность между числом правильных и неправильных решений, по оси абсцисс — номера предъявлений; Б — усредненные кривые решения задачи на оперирование эмпирической размерностью фигур и контрольного теста. По оси ординат — доля правильных выборов; по оси абсцисс — номера предъявлений
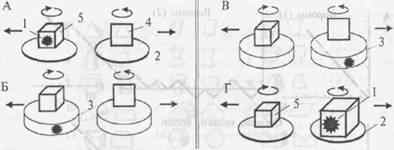
Рис. 4.17.Схемы опыта по оперированию эмпирической размерностью фигур (А), контрольных опытов по выработке дифференцировочного УР на предъявление ОФ и ПФ (Б, В) и по дифференцированию двух объемных фигур разного размера (Г) (см. текст; по Дашевскому, 1979).
В данной модификации контрольная задача теряет однозначность решения, поскольку приманка может с равной вероятностью находиться как в одной, так и в другой кормушке (тогда как в задаче на ОЭРФ она могла быть спрятана только в ОФ).
В этом варианте задачи использовали те же самые зрительные раздражители: тот же набор ОФ и ПФ, что и в задаче на ОЭРФ (рис. 4.15). У одной группы особей каждого вида (собаки, кошки, врановые) подкрепляли выбор ОФ, у другой — ПФ. В обоих случаях при первых предъявлениях животные обеих групп выбирали фигуры чисто случайно, и лишь постепенно, после десятков сочетаний они начинали чаще выбирать подкрепляемую фигуру, т.е. вырабатывали дифференцировочный УР. Следует отметить, что, как и в задаче на ОЭРФ, каждый раз животному предъявляли новую пару фигур, которая отличалась от предыдущих по всем второстепенным признакам, кроме одного: одна фигура была плоской, а другая объемной. Тем самым процедура соответствовала выработке дифференцировочного УР на обобщенный признак «размерность» (см. 3.3 и 5.5).
Как показывает рис. 4.16Б, динамика обучения дифференцировке существенно отличается от динамики решения задачи на ОЭРФ. Она сравнима с той, которая характерна для животных, плохо решающих тест на ОЭРФ (например, собак) и не имеет ничего общего с динамикой реакций у животных, хорошо справляющихся с задачей (обезьяны, дельфины, врановые птицы).
Таким путем были впервые продемонстрированы четкие различия в поведении животных при решении элементарных логических задач и при выработке дифференцировочного УР, т.е. задачи, где логическая структура отсутствует (Дашевский, Детлаф, 1974; Дашевский, 1979; Крушинский и др., 1981).
Животные, способные к решению задачи на «размерность», уже в первых предъявлениях теста реагируют правильно. При такой же по внешним признакам задаче, но требующей выработки дифференцировочного УР, правильные ответы появляются после десятков предъявлений.
Задача на поиск приманки вдвух объемных фигурах разного объема.Успешное решение теста на ОЭРФ позволило предположить, что врановым могут быть доступны и другие задачи, основанные на оперировании представлением о геометрических свойствах предметов. Для проверки этого предположения может служить тест, в котором используются две ОФ, одинаковые по форме и цвету, но существенно различающиеся по объему. За счет этого, хотя обе они обладают свойством «вмещаемости», лишь одна из них может вместить данную приманку, поскольку ее объем превышает объем кормушки в 2—4 раза, а объем второй фигуры сопоставим с ней (рис. 4.17Г).
Для решения этого теста необходимо не только качественно оценить фигуры по признаку их размерности, но произвести и количественное сопоставление их параметров. В этой связи задачу с двумя ОФ можно рассматривать как комбинированный тест, требующий оперирования сразу двумя параметрами стимулов — пространственно-геометрическими и количественными. Оказалось, что при первом предъявлении задачи птицы с равной вероятностью выбирали обе фигуры, однако при ее повторениях (от 6 до 10 раз) они достоверно чаще выбирали большую фигуру.
Опыты были проведены на 20 птицах, имевших разный опыт участия в экспериментах: 10 из них ранее успешно решили задачу на ОЭРФ, 5 птиц с этой задачей не справлялись, а еще 5 предварительно вообще не были тестированы. В их поведении при решении этой задачи, как и задачи на ОЭРФ, обнаружились значительные индивидуальные различия: 7 птиц (из 20) достоверно чаще выбирали большую ОФ (в среднем в 87% случаев); 5 птиц выбирали большую фигуру, но это предпочтение было недостоверно (примерно 65%); 4 птицы выбирали обе фигуры одинаково часто, а у 2 птиц обнаружилось предпочтение меньшей фигуры.
Эти индивидуальные особенности птиц при решении данного теста соответствовали показателям решения теста на ОЭРФ. Чем выше были они в тесте на ОЭРФ, тем легче эти птицы справлялись и с «фигурами разного объема». Однако, способность к решению основного теста на оперирование размерностью — условие необходимое, но не достаточное для решения второго.
Как уже указывалось, предполагаемый механизм решения таких тестов — мысленное сопоставление пространственных характеристик имеющихся при выборе фигур и отсутствующей в момент выбора приманки, которая служит как бы эталоном для их сопоставления (Дашевский, 1979). Опыты с использованием двух ОФ, из которых лишь одна могла вместить объемную приманку, также свидетельствуют об участии указанного механизма — мысленного сопоставления параметров фигур и отсутствующей вмомент выбора приманки. Поскольку эту задачу решает меньшая доля особей, можно заключить, что она представляет для птиц большую сложность, чем предыдущая.
Врановые птицы, дельфины, медведи и обезьяны способны к решению элементарных логических задач, основанных на оперировании пространственно-геометрическими признаками предметов.
4.7. Изучение способности животных к экстренному определению алгоритма изменений положения скрытой приманки. Тест Ревеша-Крушинского.
Этот тест был предложен Дж. Ревешем (Revecz, 1925) для сравнительной оценки рассудочной деятельности обезьян и детей, а позднее и независимо от него использовался Л. В. Крушинским, О. О. Якименко иН. П. Поповой (1983) для изучения онтогенеза невербального мышления человека. Предполагалось, что его можно рассматривать как аналог задачи на экстраполяцию, более подходящий для опытов на человеке.
Опыт ставится следующим образом. Перед животным располагают ряд одинаковых непрозрачных кормушек, накрытых крышками (испытуемым демонстрируют ряд стаканов). В первый раз приманку вне поля зрения животного помещают в первую кормушку и предоставляют возможность ее отыскать. Во второй раз (также незаметно) приманку помещают во вторую кормушку, затем в третью и т.д. После того как приманка обнаружена в первой (1-е предъявление), а затем во второй (2-е предъявление) кормушках, животное уже имеет необходимую и достаточную информацию, чтобы понять, где будет спрятана приманка при следующем предъявлении теста. Иными словами, этой информации достаточно, чтобы определить закономерность дальнейшего перемещения приманки: она каждый раз будет находиться в новом месте, ближайшем к предыдущему (рис. 4.18).
На рисунке 4.18Б видно, что галка (график слева) находила приманку безошибочно в предъявлениях с 8-го по 11-е, а в 7-м ошиблась только на один «шаг»; у павиана-анубиса (в середине) безошибочные выборы были в 5-м и 6-м. а также в 9-м и 10-м предъявлениях теста; серая крыса (справа) не сделала ни одного безошибочного выбора.
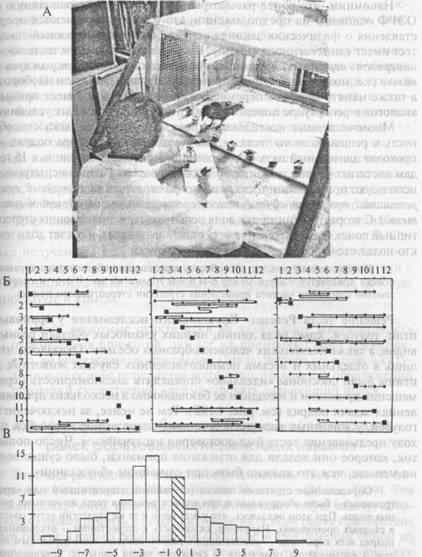
Рис. 4.18.Тест Ревеша—Крушинского.
А — обстановка эксперимента на врановых птицах в лаборатории Л. В. Крушинского; Б — результаты решения теста галкой (слева), павианом-анубисом (в середине) и серой крысой (справа); по вертикали отложены предъявления теста; по горизонтали — номера кормушек; положение приманки обозначено квадратом; неправильные реакции животного отмечены ромбами, соединенными линией; правильное (без предварительных ошибок) решение обозначено одиночным квадратом; В — гистограмма распределения первых выборов кормушек (см. текст).
Напомним, что ранее рассмотренные тесты на экстраполяцию и ОЭРФ основаны на предположении, что у животных имеются представления о физических законах окружающего мира. Каждый такой тест имеет единственное решение. В то же время в данном тесте закономерность перемещения приманки задается экспериментатором произвольно (т.е. можно перемешать приманку справа налево или наоборот, а также менять «шаг» ее перемещения). Эта задача не имеет прямых аналогов в репертуаре поведения животных в естественных условиях.
Многочисленные исследования показали, что у человека способность к решению этого
Дата добавления: 2021-09-25; просмотров: 796;
Поиск по сайту
Узнать еще
- IV. Примеры ситуационных задач
- А. Классификация видов моделирования (Л.5)
- Адаптации глубоководных видов
- Адаптации животных (особенности фауны)
- Адаптации животных к дефициту кислорода
- Адаптации особей, популяций, видов в процессе акклиматизации
- Алгоритм действий в период радиационной аварии
- Алгоритм действий при управлении автомобилем с АКПП

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
