Слизисто-бикарбонатный барьер
Слизисто–бикарбонатный барьер защищает слизистую оболочку от действия кислоты, пепсина и других потенциальных повреждающих агентов.
· Слизь постоянно секретируется на внутреннюю поверхность стенки желудка.
· Бикарбонат (ионы HCO3–), секретируемый поверхностными слизистыми клетками (рис. 22–7,I), имеет нейтрализующее действие.
· pH. Слой слизи имеет градиент pH. На поверхности слоя слизи pH равен 2, а в примембранной части более 7.
· Н+. Проницаемость плазмолеммы слизистых клеток желудка для Н+ различна. Она незначительна в мембране, обращённой в просвет органа (апикальной), и достаточно высока в базальной части. При механическом повреждении слизистой оболочки, при воздействии на неё продуктов окисления, алкоголя, слабых кислот или жёлчи концентрация H+ в клетках возрастает, что приводит к их гибели и разрушению барьера.
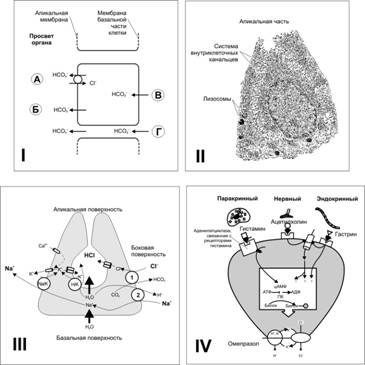
Рис. 22–7. ЖЕЛУДОЧНАЯ СЕКРЕЦИЯ. I —. Механизм секреции HCO3— эпителиальными клетками слизистой оболочки желудка и двенадцатиперстной кишки: А — выход HCO3– в обмен на Cl– стимулируют некоторые гормоны (например, глюкагон), и подавляет блокатор транспорта Cl– фуросемид. Б — активный транспорт HCO3–, не зависящий от транспорта Cl–; В и Г — транспорт HCO3– через мембрану базальной части клетки внутрь клетки и по межклеточным пространствам (зависит от гидростатического давления в подэпителиальной соединительной ткани слизистой оболочки). II — Париетальная клетка. Система внутриклеточных канальцев значительно увеличивает площадь поверхности плазматической мембраны. В многочисленных митохондриях вырабатывается АТФ для обеспечения работы ионных насосов плазматической мембраны.. III — Париетальная клетка: транспорт ионов и секреция HCl. Na+,К+‑АТФаза участвует в транспорте K+ внутрь клетки. Cl– входит в клетку в обмен на HCO3– через мембрану боковой поверхности (1), а выходит через апикальную мембрану; 2 — обмен Na+ на H+. Одно из важнейших звеньев — выход H+ через апикальную мембрану по всей поверхности внутриклеточных канальцев в обмен на K+ при помощи H+,К+‑АТФазы.. IV — Регуляции активности париетальных клеток. Стимулирующее влияние гистамина опосредуется через цАМФ, тогда как эффекты ацетилхолина и гастрина — через увеличение притока Са2+ в клетку. Простагландины снижают секрецию HCl, ингибируя аденилатциклазу, что приводит к уменьшению уровня внутриклеточного цАМФ. Блокатор H+,K+‑АТФазы (например, омепразол) снижает выработку HCl. ПК — протеинкиназа, активируемая цАМФ; фосфорилирует мембранные белки, усиливая работу ионных насосов. [11].
· Регуляция. Секрецию бикарбоната и слизи усиливают глюкагон, простагландин Е, гастрин, эпидермальный фактор роста. Для предупреждения повреждения и для восстановления повреждённого барьера применяют антисекреторные агенты (например, блокаторы гистаминовых рецепторов), простагландины, гастрин, аналоги сахаров (например, сукральфат).
· Разрушение барьера. При неблагоприятных условиях барьер разрушается в течение нескольких минут, происходят гибель клеток эпителия, отёк и кровоизлияния в собственном слое слизистой оболочки. Известны факторы, неблагоприятные для поддержания барьера: Ú нестероидные противовоспалительные препараты (например, аспирин, индометацин); Ú этанол, Ú соли жёлчных кислот, Ú Helicobacter pylori — грамотрицательная бактерия, выживающая в кислой среде желудка. H. pylori поражает поверхностный эпителий желудка и разрушает барьер, способствуя развитию гастрита и язвенного дефекта стенки желудка. Этот микроорганизм выделяют у 70% больных язвенной болезнью желудка и 90% больных язвой двенадцатиперстной кишки или антральным гастритом.
· Регенерация эпителия, формирующего слой бикарбонатной слизи, происходит за счёт стволовых клеток, расположенных на дне желудочных ямок; время обновления клеток — около 3 суток. Стимуляторы регенерации: Ú гастрин из эндокринных клеток желудка, Ú гастрин-рилизинг гормон из эндокринных клеток и окончаний волокон блуждающего нерва, Ú эпидермальный фактор роста, поступающий из слюнных, пилорических желёз, желёз двенадцатиперстной кишки и других источников.
Слизь. Помимо поверхностных клеток слизистой оболочки желудка, слизь секретируют клетки практически всех желёз желудка.
Пепсиноген. Главные клетки фундальных желёз синтезирует и секретирует предшественники пепсина (пепсиноген), а также небольшое количество липазы и амилазы. Пепсиноген не обладает пищеварительной активностью. Под влиянием соляной кислоты и особенно ранее образованного пепсина пепсиноген превращается в активный пепсин. Пепсин — протеолитический фермент, активный в кислой среде (оптимум pH от 1,8 до 3,5). При pH около 5 он практически не имеет протеолитической активности и за короткое время полностью инактивируется.
Внутренний фактор. Для всасывания витамина B12 в кишечнике необходим (внутренний) фактор Касла, синтезируемый париетальными клетками желудка. Фактор связывает витамин B12 и защищает его от разрушения ферментами. Комплекс внутреннего фактора с витамином B12 в присутствии ионов Ca2+ взаимодействует с рецепторами эпителиальной клетки дистального отдела подвздошной кишки. При этом витамин B12 поступает в клетку, а внутренний фактор высвобождается. Отсутствие внутреннего фактора приводит к развитию анемии.
Соляная кислота
Соляную кислоту (HCl) вырабатывают париетальные клетки, имеющие мощную систему внутриклеточных канальцев (рис. 22–7,II), существенно увеличивающих секреторную поверхность. Клеточная мембрана, обращённая в просвет канальцев, содержит протонный насос (H+,K+-ATФaза), выкачивающий из клетки Н+ в обмен на К+. Хлорно–бикарбонатный анионообменник встроен в мембрану боковой и базальной поверхности клеток: Cl– входит в клетку в обмен на HCO3– через этот анионообменник и выходит в просвет канальцев. Таким образом, в просвете канальцев оказываются оба компонента соляной кислоты: и Cl–, и H+. Все остальные молекулярные компоненты (ферменты, ионные насосы, трансмембранные переносчики) направлены на сохранение ионного баланса внутри клетки, прежде всего — на поддержание внутриклеточного pH.
Таким образом, механизм образования соляной кислоты (рис. 22–7,II) складывается из следующих этапов.
· Ионы хлора активно транспортируются из цитоплазмы обкладочных клеток в просвет внутриклеточных канальцев, а ионы натрия переносятся наружу. Эти два эффекта создают в канальцах потенциал (от –40 до –70 мВ), приводящий к диффузии большого количества ионов калия и некоторого количества ионов натрия внутрь канальцев.
· В цитоплазме клетки вода диссоциирует на водородные и гидроксильные ионы. Водородные ионы посредством активного транспорта секретируются внутрь канальцев в обмен на ионы калия. Этот процесс катализирует H+, K+‑АТФаза. В результате ионы калия и натрия, которые диффундировали внутрь канальцев, реабсорбируются в цитоплазму клеток, а ионы водорода занимают их место в канальцах, создавая там раствор концентрированной соляной кислоты, который через секреторные канальцы поступает наружу, т.е. на поверхность эпителия желудка.
· Вода проходит внутрь канальцев под воздействием осмоса. В результате секрет канальцев содержит соляную кислоту в концентрации 150–160 ммоль/л, хлорид калия в концентрации 15 ммоль/л и небольшое количество хлорида натрия.
· На заключительным этапе углекислота, образующаяся в ходе метаболизма клетки, под влиянием карбоангидразы соединяется с гидроксильными ионами, образуя бикарбонатные ионы. Бикарбонатные ионы диффундируют из цитоплазмы во внеклеточную жидкость в обмен на ионы хлора, входящие в клетку и в дальнейшем секретируемые в канальцы париетальных клеток.
· В покое клетка секретирует хлор.
· При стимуляции усиливается работа Н+,К+‑АТФазы, в результате чего возрастает транспорт Н+ из клетки. Одновременно в 2–3 раза увеличивается транспорт Cl– в обмен на HCO3–. В итоге внутриклеточный pH остаётся неизменным. Стимуляция обкладочных клеток вызывает выделение кислого раствора, изотоничного внеклеточной жидкости, содержащего 160 ммоль/л соляной кислоты с величиной pH раствора около 0,8. Содержание водородных ионов в этом растворе в 3 млн раз выше, чем в артериальной крови.
Регуляция секреции соляной кислоты представлена на рис. 22–7,IV. Париетальная клетка активируется через м‑холинорецепторы (блокатор — атропин), Н2-рецепторы гистамина (блокатор — циметидин) и рецепторы гастрина (блокатор — проглюмид). Указанные блокаторы или их аналоги, а также ваготомия используются для подавления секреции соляной кислоты. Существует ещё один способ понизить выработку соляной кислоты — блокада Н+,К+‑АТФазы.
Дата добавления: 2016-06-05; просмотров: 3366;
Поиск по сайту
Узнать еще
- II. РАСПРЕДЕЛЕНИЕ ЛЕКАРСТВЕННЫХ СРЕДСТВ В ОРГАНИЗМЕ. БИОЛОГИЧЕСКИЕ БАРЬЕРЫ. ДЕПОНИРОВАНИЕ
- АГБ – аэрогематический барьер
- Барьер психической адаптации и этиопатогенетическая сущность пограничных состояний
- Барьерная емкость р-n-перехода
- Барьерное заводнение
- БАРЬЕРЫ В КОММУНИКАЦИИ
- Барьеры входа на рынок и выхода с рынка как показатели структуры рынка. Нестратегические барьеры
- Барьеры общения и их преодоление

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
