Мейоз как основа полового размножения
Мейозом называется особый способ деления эукариотических клеток, при котором исходное число хромосом уменьшается в 2 раза (от древнегреч. «мейон» – меньше – и от «мейозис» – уменьшение).
Отдельные фазы мейоза у животных описал В. Флемминг (1882), а у растений – Э.Страсбургер (1888), а затем российский ученый В.И. Беляев. В это же время (1887) А. Вайсман теоретически обосновал необходимость мейоза как механизма поддержания постоянного числа хромосом. Первое подробное описание мейоза в ооцитах кролика дал Уиниуортер (1900).
Хотя мейоз открыт более 100 лет назад, но изучение мейоза продолжается до сих пор. Интерес к мейозу резко возрос в конце 60-х годов, когда выяснилось, что одни и те же контролируемые генами ферменты могут принимать участие во многих процессах, связанных с ДНК. В последнее время ряд биологов развивают оригинальную идею: мейоз у высших организмов служит гарантом стабильности генетического материала, ибо в процессе мейоза, когда пары хромосом-гомологов тесно соприкасаются, происходит проверка нитей ДНК на точность и восстановление повреждений, затрагивающих сразу обе нити. Изучение мейоза связало методы и интересы двух наук: цитологии и генетики. Это привело к рождению новой ветки знания — цитогенетики, тесно соприкасающейся ныне с молекулярной биологией и генной инженерией.
Биологическое значение мейоза заключается в следующих процессах:
1.Благодаря редукции числа хромосом в результате мейоза в ряду поколений при половом размножении обеспечивается постоянство числа хромосом.
2.Независимое распределение хромосом в анафазе первого деления обеспечивает рекомбинацию генов, относящихся к разным группам сцепления (находящихся в разных хромосомах). Мейотическое распределение хромосом по дочерним клеткам называется сегрегацией хромосом.
3.Кроссинговер в профазе I мейоза обеспечивает рекомбинацию генов, относящихся к одной группе сцепления (находящихся в одной хромосоме).
4. Случайное сочетание гамет при оплодотворении вместе с вышеперечисленными процессами способствует генетической изменчивости.
5. В процессе мейоза происходит еще одно существенное явление. Это процесс активации синтеза РНК (или транскрипционной деятельности хромосом) в ходе профазы (диплотены), связанный с формированием хромосом типа «ламповых щеток» (обнаружены у животных и некоторых растений).
Эта реверсия профазы к интерфазному состоянию (при митозе только в интерфазе идет синтез и-РНК) является специфической характеристикой мейоза как особого типа деления клеток.
Следует отметить, что у простейших наблюдается значительное разнообразие процессов мейоза.
В соответствии с положением в жизненном цикле различают три типа мейоза:
Зиготный (исходный) мейоз происходит в зиготе, т.е. непосредственно после оплодотворения. Он характерен для организмов, в жизненном цикле которых преобладает гаплоидная фаза (аскомицеты, бизидиомицеты, некоторые водоросли, споровики и др.).
Гаметный (терминальный) мейоз происходит во время формирования гамет. Он наблюдается у многоклеточных животных (в т.ч. у человека), а также среди простейших и некоторых низших растений, в жизненном цикле которых преобладает диплоидная фаза.
Промежуточный (споровый) мейоз протекает во время спорообразования у высших растений, включаясь между стадиями спорофита (растения) и гаметофита (пыльца, зародышевый мешок).
Таким образом, мейоз - это форма ядерного деления, сопровождающаяся уменьшением числа хромосом с диплоидного до гаплоидного и изменением генетического материала. Результат мейоза — образование клеток с гаплоидным набором хромосом (половых клеток).
Продолжительность мейоза может отличаться в зависимости от вида растений и животных (табл. 1).
Таблица 1. Продолжительность мейоза у различных видов растений
| Вид | 2n | Мейотический цикл, час |
| Secate sereate (рожь культурная) | 51,2 | |
| Allium сера (лук репчатый) | 96,0 | |
| Tradescantia paludosa | 126,0 | |
| Lilium herry | 170,0 | |
| Lilium longiflorum | 192,0 | |
| Trillium erectum (трилиум прямостоячий) | 274,0 |
Типичный мейоз состоит из двух последовательных клеточных делений, которые соответственно называются мейоз I и мейоз II. В первом делении происходит уменьшение числа хромосом в два раза, поэтому первое мейотическое деление называют редукционным, реже – гетеротипным. Во втором делении число хромосом не изменяется; такое деление называют эквационным (уравнивающим), реже – гомеотипным. Выражения «мейоз» и «редукционное деление» часто используют как синонимы.
Исходное число хромосом в мейоцитах (клетках, вступающих в мейоз) называется диплоидным хромосомным числом (2n) Число хромосом в клетках, образовавшихся в результате мейоза, называется гаплоидным хромосомным числом (n). Минимальное число хромосом в клетке называется основным числом (x). Основному числу хромосом в клетке соответствует и минимальный объем генетической информации (минимальный объем ДНК), который называется геном.
Количество геномов в клетке называется геномным числом (n). У большинства многоклеточных животных, у всех голосеменных и многих покрытосеменных растений понятие гаплоидности–диплоидности и понятие геномного числа совпадают. Например, у человека n=x=23 и 2n=2x=46.
Морфология мейоза — характеристика фаз
Интерфаза
Предмейотическая интерфаза отличается от обычной интерфазы тем, что процесс репликации ДНК не доходит до конца: примерно 0,2...0,4 % ДНК остается неудвоенной. Таким образом, деление клетки начинается на синтетической стадии клеточного цикла. Поэтому мейоз образно называют преждевременным митозом. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с.
При наличии центриолей происходит их удвоение таким образом, что в клетке имеется две диплосомы, каждая из которых содержит пару центриолей.
Первое деление мейоза
ДНК прошла репликацию. Начитается профаза I, самая продолжительная стадия мейоза.
Стадия профазы I подразделяется на следующие стадии:
лептотена — стадия тонких нитей;
зиготена — стадия двойных нитей;
пахитена — стадия толстых нитей;
диплотена — кроссинговер;
диакинез — исчезновение ядерной оболочки и ядрышка.
В ранней профазе (лептотене) происходит подготовка к конъюгации хромосом. Хромосомы уже удвоены, но сестринские хроматиды в них еще неразличимы. Хромосомы начинают упаковываться (спирализоваться).
В отличие от профазы митоза, где хромосомы расположены по мембране ядра конец в конец и, упаковываясь, притягиваются к мембране, лептотенные хромосомы своими теломерными участками (концами) располагаются в одном из полюсов ядра, образуя фигуру «букета» у животных и сжатие в клубок «синезис» — у растений. Такое расположение или ориентации в ядре позволяет хромосомам быстрее и легче осуществлять конъюгацию гомологичных локусов хромосом (рис. 1).
Центральное событие — таинственный процесс узнавания гомологичных хромосом и их попарное сближение друг с другом происходит в зиготене профазы I. При конъюгации (сближении) гомологичных хромосом происходит образование пар — бивалентов и хромосомы заметно укорачиваются. С этого момента начинается формирование синаптонемного комплекса (СК). Формирование синаптонемного комплекса и синопсис хромосом — синонимы.
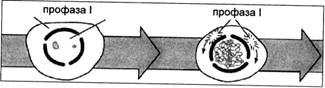
Рис. 1. Стадия профазы
В ходе следующей стадии профазы I – пахитене между гомологичными хромосомами усивается тесное соприкосновение, которое и называется синапсисом (от греч. synopsis — соединение, связь). Хромосомы в этой стадии сильно спирализованы, что делает возможным наблюдение их под микроскопом.
В ходе синапсиса гомологи переплетаются, т.е. конъюгируют. Конъюгирующие биваленты связаны хиазмами. Каждый бивалент состоит из двух хромосом и четырех хроматид, где каждая хромосома пришла от своего родителя. При образовании синапсиса (СК), происходит обмен участками между гомологичными хроматидами. Этот процесс, называемый кроссинговером, приводит к тому, что хроматиды теперь имеют иной состав генов.
Синаптонемный комплекс (СК) в пахитене достигает наибольшего развития и в этот период представляет собой лентовидную структуру, располагающуюся в пространстве между параллельно лежащими гомологичными хромосомами. СК состоит из двух параллельных латеральных элементов, сформированных плотно уложенными белками и менее плотного центрального элемента, протягивающегося между ними (рис. 2).
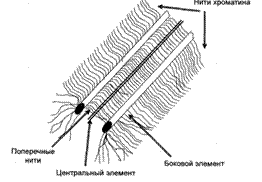
Рис. 2. Схема синаптонемного комлекса
Каждый латеральный элемент формируется парой сестринских хроматид в виде продольной оси лептотенной хромосомы и до того, как становится частью СК, носит название осевого элемента. Боковые петли хроматина лежат вне СК, окружая его со всех сторон.
Развитие СК в процессе мейоза:
лептотена—структура хромосом, вступивших в лептотену, сразу же оказывается необычной: в каждом гомологе наблюдается продольный тяж, идущий по оси хромосом на всем ее протяжении;
зиготена — на этой стадии осевые тяжи гомологов сближаются, при этом концы осевых тяжей, прикрепленных к ядерной мембране, как бы скользят по ее внутренней поверхности навстречу друг к другу;
пахитена. Наибольшее развитие СК достигает в пахитене, когда все элементы его приобретают максимальную плотность, а хроматин — вид плотной сплошной «шубы» вокруг него.
Функции СК:
1.Полностью развитый синаптонемный комплекс необходим для нормального удержания гомологов в биваленте так долго, как это необходимо для осуществления кроссинговера и закладки хиазм. Хромосомы соединяются с помощью синаптонемного комплекса на некоторое время (от 2 ч у дрожжей до 2–3 сут. у человека), в течение которого между гомологичными хромосомами совершается обмен гомологичными участками ДНК — кроссинговер (от англ, crossing over — образование перекреста).
2.Предотвращение слишком прочного соединения гомологов и удержание их на определенном расстоянии, сохранение их индивидуальности, создание возможности оттолкнуться в диплотене и разойтись в анафазе.
Процесс кроссинговера связан с работой определенных ферментов, которые при образовании хиазм между сестринскими хроматидами, «разрезают» их в месте перекреста с последующим воссоединением образовавшихся фрагментов. В большинстве случаев указанные процессы не приводят к каким-либо нарушениям в генетической структуре гомологичных хромосом, т.е. происходит правильное соединение фрагментов хроматид и восстановление их первоначального строения.
Однако, возможен и другой (более редкий) вариант событий, который связан с ошибочным воссоединением фрагментов разрезанных структур. При этом происходит взаимный обмен участками генетического материала между конъюгирующими хроматидами (генетическая рекомбинация).
На рис. 3 приведена упрощенная схема некоторых возможных вариантов одиночного либо двойного кроссинговера с участием двух хроматид из пары гомологичных хромосом. Необходимо подчеркнуть, что кроссинговер представляет собой случайное событие, которое с той или иной вероятностью может возникнуть на любом участке (либо на двух и большем числе участков) гомологичных хромосом. Следовательно, на этапе созревания гамет эукариотического организма в профазе первого деления мейоза действует универсальный принцип случайного (свободного) комбинирования (рекомбинации) генетического материала гомологичных хромосом.
В цитологических исследованиях синапсиса в последние два десятилетия важную роль играет метод распластывания профазных мейотических клеток животных и растений под действием гипотонического раствора. Метод вошел в цитогенетику после работ Мозеса и сыграл такую же роль, какую в свое время сыграл метод приготовления «давленых» препаратов для исследования метафазных хромосом, избавив цитогенетиков от микротомных срезов.
Метод Мозеса и его модификации стали более удобными, чем анализ СК на ультратонких срезах. Этот метод был положен в основу исследований мейоза и постепенно охватил вопросы генного контроля мейоза у животных и растений.
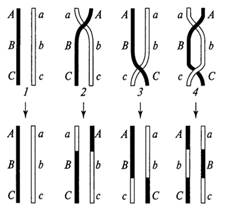
Рис. 3. Отдельные варианты одиночного и двойного кроссинговера с участием двух хроматид: 1 исходные хроматиды и вариант без кроссинговера; 2 одиночный кроссинговер на участке А В и кроссоверные хроматиды; 3 одиночный кроссинговер на участке В-С и кроссоверные хроматиды; 4 двойной кроссинговер и кроссоверные хроматиды нескольких разных участках на основе гомологичности генетического материала этих участков. Полагают, что с каждой стороны в процессе конъюгации могут участвовать либо одна из двух сестринских хроматид соответствующей хромосомы либо обе хроматиды.
В диппотене гомологичные хромосомы после спаривания и кроссинговера начинают отталкиваться друг от друга. Процесс отталкивания начинается с центромер. Расхождению гомологов препятствуют хиазмы - место соединения несестринских хроматид, возникших в результате перекреста. По мере расхождения хроматид некоторые хиазмы смещаются к концу плеча хромосомы. Обычно перекрестов бывает несколько, и чем длиннее хромосомы, тем их больше, поэтому в диплотене, как правило, несколько хиазм в одном биваленте.
В стадии диакинеза происходит уменьшение числа хиазм. Биваленты располагаются по периферии ядра. Ядрышко растворяется, мембрана разрушается и начинается переход к метафазе I. На протяжении всей профазы сохраняется ядрышко и ядерная оболочка. Перед профазой в период синтетического периода интерфазы происходит репликация ДНК и репродукция хромосом. Однако полностью этот синтез не заканчивается: ДНК синтезируется на 99,8%, а белки — на 75%. Синтез ДНК заканчивается в пахитене, белков — в диплотене.
В метафазе I становится заметной веретеновидная структура, образуемая микротрубочками. В ходе мейоза к центромерам хромосом каждого бивалента прикрепляются отдельные микрокрубочки. Затем пары хромосом перемещаются в экваториальную плоскость клетки, где выстраиваются в случайном порядке. Центромеры гомологичных хромосом располагаются в противоположных сторонах от экваториальной плоскости; в метафазе митоза, напротив, центромеры отдельных хромосом располагаются в экваториальной плоскости.
В метафазе I биваленты располагаются в центре клетки, в зоне экваториальной пластинки (рис. 4).
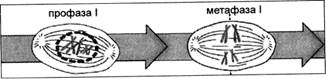
Рис. 4. Стадии мейоза: профаза I — метафаза I
Анафаза начинается с расхождения гомологичных хромосом и движения их в направлении полюсов. У хромосом без центромера крепления не может существовать. В анафазе митоза центромеры делятся и идентичные хроматиды расходятся. В анафазе I мейоза центромеры не делятся, хроматиды остаются вместе, а разъединяются гомологичные хромосомы. Однако из-за обмена фрагментами в результате кроссинговера хроматиды не идентичны, как в начале мейоза. В анафазе I конъюгирующие гомологи расходятся к полюсам.
В дочерних клетках число хромосом вдвое меньше (гаплоидный набор), при этом масса ДНК уменьшается также вдвое и хромосомы остаются дихроматидными. Точное расхождение гомологичных пар к противоположным полюсам лежит в основе редукции их числа.
В телофазе I происходит сосредоточение хромосом у полюсов, некоторая их деконденсация, за счет чего спирализация хромосом ослабевает, они удлиняются и снова становятся неразличимыми (рис. 5). По мере того как телофаза постепенно переходит в интерфазу, из эндоплазматического ретикулума возникает ядерная оболочка (в том числе и из фрагментов оболочки ядра материнской клетки), а также клеточная перегородка. Наконец вновь образуется ядрышко и возобновляется синтез белка.
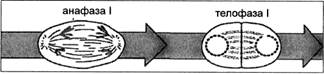
Рис. 5. Стадии мейоза: анафаза I — телофаза I
В интеркинезе образуются ядра, в каждой из которых находится n дихроматидных хромосом.
Особенность второго деления мейоза состоит, прежде всего, в том, что в интерфазе II не происходит удвоения хроматина, поэтому каждая клетка, вступающая в профазу II, сохраняет прежнее соотношение n2с.
Второе деление мейоза
В период второго деления мейоза сестринские хроматиды каждой хромосомы расходятся к полюсам. Поскольку в профазе I мог произойти кроссинговер и сестринские хроматиды могли стать неидентичными, то принято говорить, что второе деление протекает по типу митоза, однако это не настоящий митоз, при котором в норме дочерние клетки содержат хромосомы идентичные по форме и набору генов.
В начале второго мейотического деления хроматиды все еще связаны центромерами. Это деление похоже на митоз: если в телофазе I образовалась ядерная оболочка, то теперь она разрушается, и к концу короткой профазы II исчезает ядрышко.
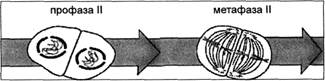
Рис. 6. Стадии мейоза: профаза II—метафаза II
В метафазе II снова можно увидеть веретено и хромосомы, состоящие из двух хроматид. Хромосомы прикрепляются центромерами к нитям веретена и выстраиваются в экваториальной плоскости (рис. 6). В анафазе II центромеры делятся и расходятся, а сестринские хроматиды, ставшие теперь хромосомами, движутся к противоположным полюсам. В телофазе II образуются новые ядерные оболочки и ядрышки, сжатие хромосом ослабевает и в интерфазном ядре они становятся невидимыми (рис. 7).
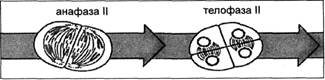
Рис. 7. Стадии мейоза: анафаза II — телофаза II
Завершается мейоз формированием гаплоидных клеток — гаметы, тетрады спор - потомков исходной клетки с редукционным вдвое (гаплоидным) набором хромосом и гаплоидной массой ДНК (исходная клетка 2n, 4с, — споры, гаметы — n, с).
Общая схема распределения хромосом гомологичной пары и содержащихся в них двух пар различающихся аллельных генов во время двух делений мейоза приведена на рис.8. Как видно из этой схемы, возможны два принципиально разных варианта такого распределения. Первый (более вероятный) вариант связан с образованием двух типов генетически различающихся гамет с хромосомами, не претерпевшими кроссинговеров на участках, где локализованы рассматриваемые гены. Такие гаметы принято называть некроссоверными. При втором (менее вероятном) варианте наряду с некроссоверными возникают также кроссоверные гаметы как результат генетического обмена (генетической рекомбинации) в участках гомологичных хромосом, расположенных между локусами двух неаллельных генов.
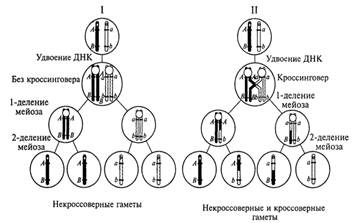
Рис. 8. Два варианта распределения хромосом гомологичной пары и содержащихся в них неаллельных генов как результат двух делений мейоза
Дата добавления: 2017-03-12; просмотров: 5302;
Поиск по сайту
Узнать еще
- I. Демократия как тип политического режима.
- I. Судовождение, основанное только на лоцманском методе.
- II. Агрессия как проявление побуждения
- II. КРОВЬ КАК ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА. ФИЗИОЛОГИЯ ЭРИТРОЦИТОВ И ЛЕЙКОЦИТОВ.
- Windows 9X как пример системы, управляемой событиями
- А - при выпучине; б - при просадке основания; 1 - выпучина; 2 - отверстие; 3 -гидрофобный грунт; 4 - наладка; 5 - просадка основания; 6 - сварной шов; 7 -днище.
- Абсолютный показатель базисный (база сравнения, основание)
- Авангардизм как стадия развития искусства.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
