То такое раздражимость
Взаимодействие организма с окружающей средой проявляется в форме раздражимости. Раздражимость – способность организмов (клеток) отвечать на действия раздражителей структурно-функциональными изменениями.
Раздражители – различные факторы внешней или внутренней среды, воздействующие на организм. Можно выделить физические (механические, температурные, электрические и т.п.), химические (кислоты, щелочи, соли, гормоны, медиаторы и др.), физико-химические (изменения осмотического давления, рН и др.), биологические (бактерии, вирусы и др.), информационные раздражители. Так же выделяют адекватные и неадекватные раздражители. Первые действуют на биологические объекты в естественных условиях (организмы генетически приспособлены к действию этих факторов). Вторые действуют на клетки или организм, как правило, в условиях эксперимента. Например, мышца может сокращаться как при действии медиатора (адекватный раздражитель), так и при действии электрического тока. Можно разделять раздражители на пороговые, подпороговые или сверхпороговые (смотрите лабораторные работы!!).
Активно взаимодействуя с разнообразными раздражителями внешней среды биологические организмы отражали в своей организации значимые события окружающего мира (приспосабливались, адаптировались к этим событиям).
Нейроны, мышечная ткань и клетки секреторного эпителия кроме раздражимости обладают еще и возбудимостью. Возбудимость – способность клеток отвечать на действие раздражителей изменением мембранного потенциала.
В начале XX века установлено, что между внешней и внутренней сторонами мембраны клеток существует разность электрических потенциалов. Ю.Бернштейн впервые назвал эту разность мембранным потенциалом, возникающим в связи с неравномерным распределением ионов по обе стороны цитоплазматической мембраны.
В 30-40-х годах XX века был открыт гигантский аксон кальмара диаметром 0.5 мм (Янг; 1936) и изготовлен стеклянный микроэлектрод с диаметром кончика около 1 мкм (Грехам, Герард; 1946). Эти открытия позволили Ходжкину и Хаксли измерить электрическую разность потенциалов между аксоплазмой и наружной средой.Таким образом было установлено, что разница потенциалов по обе стороны мембраны клеток возбудимых тканей составляет определенную величину: от -50 до -100 мВ. Это означает, что внутри мембрана заряжена отрицательно, а снаружи – положительно(рис. 1).
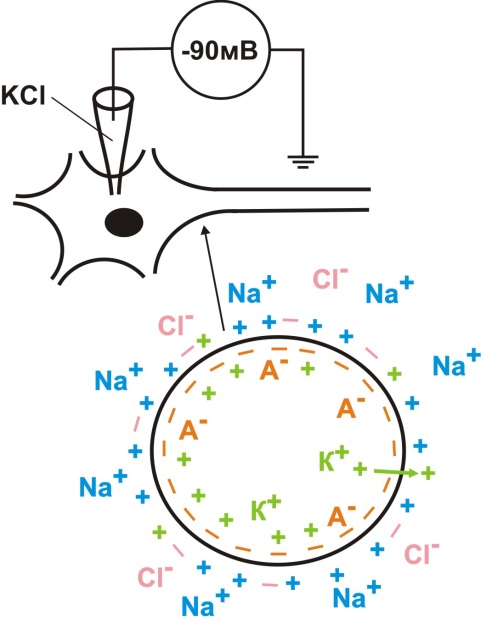
Рис. 1. Схема регистрации мембранного потенциала
Отрицательный заряд внутри обусловлен суммарным зарядом белков-анионов (крупные белковые молекулы не могут покинуть клетку через неповрежденную мембрану). Положительный заряд снаружи обусловлен суммарным зарядом ионов Na+. Оказалось, что цитоплазма и окружающая клетку среда содержат разное количество ионов К+, Na+, Ca2+, Cl- (таблица 1).
Таблица 1
Различие концентраций ионов в цитоплазме и межклеточном веществе
| ионы | концентрация ионов, ммоль/л | ||
| внутриклеточная [in] | внеклеточная [out] | [ионin]/[ионout] | |
| Na+ | 1:9 | ||
| К+ | 20:1 | ||
| Са2+ | 0.04 | 1:25 | |
| Cl– | 1:11 | ||
| А– | – | – |
Разница концентраций каждого иона создает силу (градиент концентрации), которая обеспечивает диффузию этого иона по принципу «оттуда, где больше – туда, где меньше». В отсутствие раздражителей мембрана практически непроницаема для Na+, в то время как ионы К+ по градиенту концентрации могут выходить из клетки через мембранные каналы (так называемый ток утечки). Как долго калий будет выходить из клетки? Отношение концентраций внеклеточного К+ к внутриклеточному составляет 1/20. Однако в результате диффузного выхода калия по градиенту концентрации происходит перенос заряда катиона на внешнюю, положительно заряженную сторону мембраны. По закону электрофизики одноименные заряды отталкиваются, а разноименные - притягиваются. Значит, выход калия будет продолжаться до тех пор, пока сила концентрационного градиента не уравняется с противоположенной силой электрохимического градиента (рис. 2).
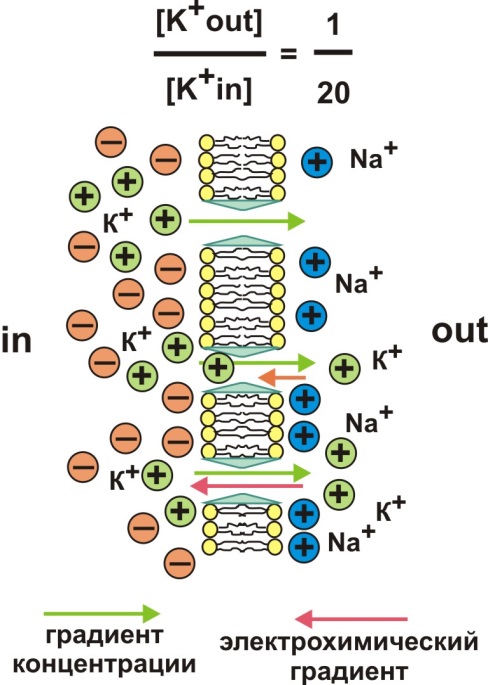
Рис. 2. Калиевый равновесный потенциал
В результате установится относительное равновесие в распределении ионов калия (и других зарядов) по обе стороны мембраны. В этих условиях электрический потенциал мембраны получил название калиевого равновесного потенциала (ЕК+). Обратите внимание, что величина данного потенциала в основном определяется разницей концентрации ионов К+ по обе стороны цитоплазматической мембраны. Равновесный калиевый потенциал можно рассчитать по формуле Нернста:
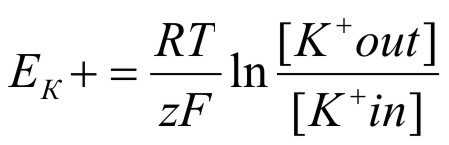
где R – газовая постоянная Больцмана; T – температура в Кельвинах; z – валентность иона; F – число Фарадея.
При постоянных условиях внешней среды левая часть формулы представляет собой константную величину и равновесный калиевый потенциал зависит только от разницы концентраций ионов калия по обе стороны мембраны.В отсутствие раздражителей равновесный калиевый потенциал относительно постоянный и носит название потенциала покоя.
Другие ионы (Na+, Ca2+, Cl-) в нормальных условиях и при отсутствии раздражителей по разным причинам существенно не влияют на величину мембранного потенциала. Поэтому можно сказать, что величина потенциала покоя в основном определяется разницей концентраций ионов калия по обе стороны цитоплазматической мембраны. От величины потенциала покоя зависит степень возбудимости мембраны. Изменение потенциала покоя может привести к нарушению процессов возбуждения. В организме 98% К+ находится внутриклеточно, однако с мочой, потом и слезной жидкостью К+ регулярно выводится из организма. Суточная потребность калия у человека составляет 2-3 г. Недостаток К+ приводит к нарушению функции сердца (аритмия, нарушение сократительной деятельности) и ЦНС (например, галлюцинации и обмороки в условиях минерального голодания или нарушения водно-солевого баланса от жары).
Таким образом мембранный потенциал в отсутствии раздражителей остается относительно постоянным. При действии раздражителей мембранный потенциал изменяется за счет изменения ионной проводимости мембраны.Представим этот процесс на схеме, отразив шкалу величины мембранного потенциала, действие раздражителя отметим стрелкой (рис. 3).
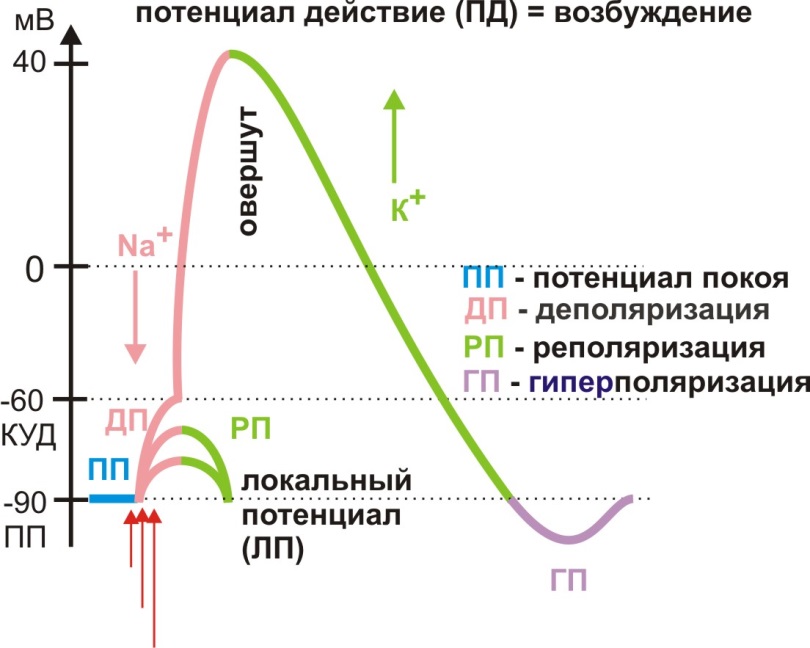
Рис. 3. Мембранные потенциалы
При действии небольшого раздражителя будет происходить уменьшение мембранного потенциала (снижение поляризации мембраны, уменьшение ее электроотрицательности). Этот процесс получил название деполяризации. Затем последует восстановление мембранного потенциала до исходной величины (ПП) – реполяризация. С увеличением силы раздражителя деполяризация развивается быстрее и на большую величину (рис. 3). Однако разницы между этими потенциалами нет – это локальные потенциалы.
Если раздражитель вызовет деполяризацию примерно на 30 мВ – до критического уровня (КУД - критический уровень деполяризации), вслед за медленной деполяризацией произойдет быстрое изменение мембранного потенциала до величины 30-40 мВ с последующей реполяризацией. Такое изменение мембранного потенциала носит название потенциала действия (ПД), в этом процессе проявляется возбуждение мембраны (рис. 3).
Процесс деполяризации обусловлен входом внутрь клетки Na+ по градиенту концентрации иона через отрывшиеся под действием раздражителя специфические натриевые мембранные каналы.Разница амплитуды ЛП и ПД объясняется количеством открывающихся натриевых каналов. Натриевый канал представляет собой мембранный канальный белок, имеющий пору, соответствующую диаметру гидратированного иона Na+, и две субъединицы, так называемые активационные и инактивационные ворота. Если начало деполяризации вызвано открытием натриевого канала (открываются активационные ворота), то завершение деполяризации связано закрытием ионного канала (закрываются инактивационные ворота). Состояние канала определяется величиной мембранного потенциала, поэтому Na+ каналявляется потенциалзависимым.
Смена внутреннего заряда мембраны с отрицательного на положительный в процессе деполяризации носит название овершута (рис. 3).
Процесс реполяризации обусловлен выходом из клетки К+ по градиенту концентрации иона через отрывшиеся специфические калиевые мембранные каналы.Калиевый канал имеет одну воротную субъединицу, более инертен, поэтому при частых возбуждениях не способен сразу закрыться, чем объясняется явление следовой гиперполяризации(рис. 3).
Длительность ПД составляет 1-2 мс, нервная клетка способна возбуждаться с частотой до 200 имп/с.
Для поддержания градиентов концентрации ионов Na+ и К+ в мембране расположена ферментная транспортная система - Na+/К+-АТФаза, с помощью которой вошедший в клетку во время деполяризации Na+ перемещается во внешнюю среду, а вышедший из клетки в процессе реполяризации К+ возвращается обратно в цитоплазму (рис. 4).
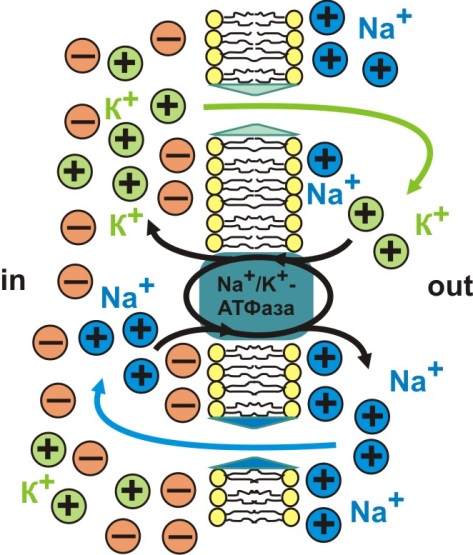
Рис. 4. Схема функционирования Na+/К+-АТФазы
Для транспорта ионов против градиента концентрации Na+/К+-АТФаза использует энергию АТФ.
Таким образом, процессы возбуждения обеспечивает работа трех транспортных систем: потенциалзависимых и избирательных Na+ и К+ каналов, а также Na+/К+-АТФазы, краткая характеристика которых представлена в таблице 2.
Таблица 2
Сравнительная характеристика транспортных систем цитоплазматической мембраны
| Свойство | Na+ канал | К+ канал | Na+/К+-АТФаза |
| Селективность транспорта | только Na+ | только К+ | сопряженный 3 Na+ меняет на 2 К+ |
| Направление транспорта | По градиенту концентрации Na+ | По градиенту концентрации К+ | Против градиента концентрации ионов |
| Энергозависимость | Нет, диффузия | Нет, диффузия | Да, АТФ |
| Потенциалзависимость | Да | Да | Нет |
| Зависимость от физико-химических факторов (рН, t) | Независим | Независим | Чувствительна к изменениям рН и t |
| Ингибиторы (блокаторы) | Тетродотоксин (рыба фуга) | Тетраэтиламмоний | Строфантин (ландыш майский) |
Мерой возбудимости является минимальная сила раздражителя, вызывающая возбуждение.Такой раздражитель называется пороговым, а его величина - порогом. Между порогом и уровнем возбудимости существует обратная зависимость: чем больше порог, тем ниже возбудимость, и наоборот. Раздражитель силой меньше порогового называют подпороговым, а больше – надпороговым. Надпороговый раздражитель, по вышеуказанному определению, несомненно, вызовет возбуждение. При этом амплитуда потенциала действия с увеличением силы надпорогового раздражителя возрастать не будет. Эта особенность носит названия закона ВСЕ ИЛИ НИЧЕГО: амплитуда потенциала действия при возбуждении максимальна. Иными словами, мембрана нейрона отражает свойства раздражителя в двоичном коде: либо в форме локального потенциала (т.е. не отвечает возбуждением если раздражитель подпороговый), либо генерирует потенциал действия (т.е. возбуждается если раздражитель пороговой или надпороговой силы).
Однако возбудимость не есть величина постоянная; даже в процессе возбуждения чувствительность мембраны к раздражителям существенно меняется. Для оценки уровня возбудимости используют три определения: нормальная возбудимость, экзальтация (повышенная возбудимость) и рефрактерность (пониженная возбудимость или невозбудимость). Если мы обозначим возбудимость мембраны в исходном состоянии покоя как нормальную, то в условиях быстрой деполяризации и начале реполяризации мембрана будет абсолютно рефрактерна (АР), т.е. невозбудима. В процессе реполяризации мембранного потенциала до величины, соответствующей КУД, мембрана будет относительно рефрактерна (ОР) и ее снова можно перевести в состояние возбуждения только надпороговым раздражителем. Когда в процессе реполяризации мембранный потенциал будет соответствовать интервалу от ПП до КУД, возбудимость мембраны будет повышенной (экзальтация мембраны). И наконец, в условиях следовой гиперполяризации возбудимость мембраны снова станет пониженной (рис. 5).
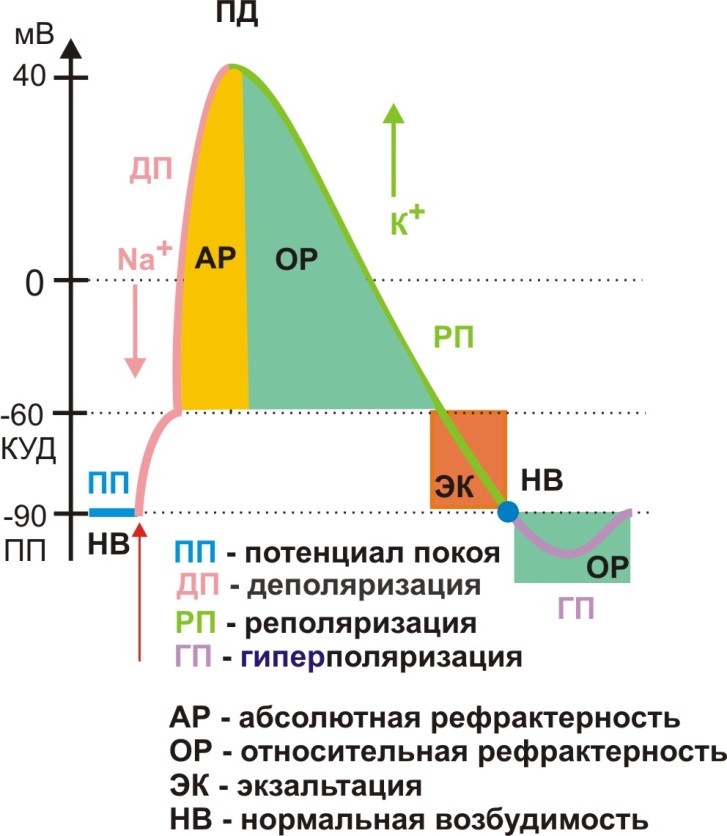
Рис. 5. Изменение возбудимости мембраны в процессе возбуждения
Длительность периода АР будет определять важное свойство возбудимых клеток: их лабильность, как способность отвечать возбуждением на действие порогового (или надпорогового) раздражителя в точном соответствии с заданным ритмом раздражения (Н.Е. Введенский, 1982). Для нервной системы в целом лабильность будет определять индивидуальную способность организма воспринимать и перерабатывать информацию с определенной скоростью. Состояние ОР или АР объясняется частичной или полной инактивацией Na+ каналов. Инактивация Na+ каналов (АР) может наступить в случае длительного воздействия на мембрану подпорогового раздражителя, что носит название аккомодации.
Важнейшими характеристиками любого раздражителя являются его амплитуда (сила) и длительность (время воздействия заданной амплитудой). По этим критериям пороговые раздражители могут отличаться друг от друга. Например, короткий импульсный пороговый раздражитель должен иметь большую силу, в тоже время возбуждение может вызвать раздражитель меньшей силы, но тогда он должен действовать более продолжительное время (рис. 6).
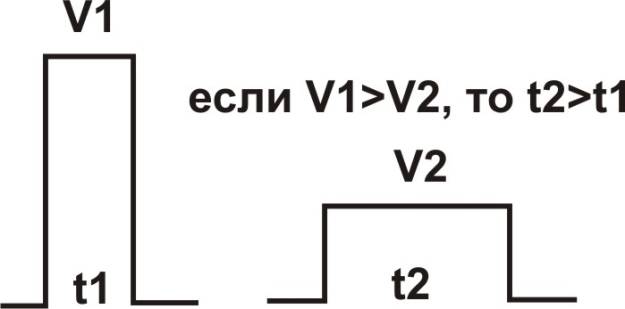
Рис. 6. Сравнительная характеристика пороговых раздражителей (V1 и V2 - амплитуда раздражителей; t1 и t2 - минимальное достаточное для возбуждения время действия раздражителей)
Зависимость силы порогового раздражителя от его длительности носит название кривой «сила-длительность» Вейса-Лапика (рис. 7). Все точки этой кривой отражают параметры порога, правее и выше кривой – параметры надпорогового раздражителя.
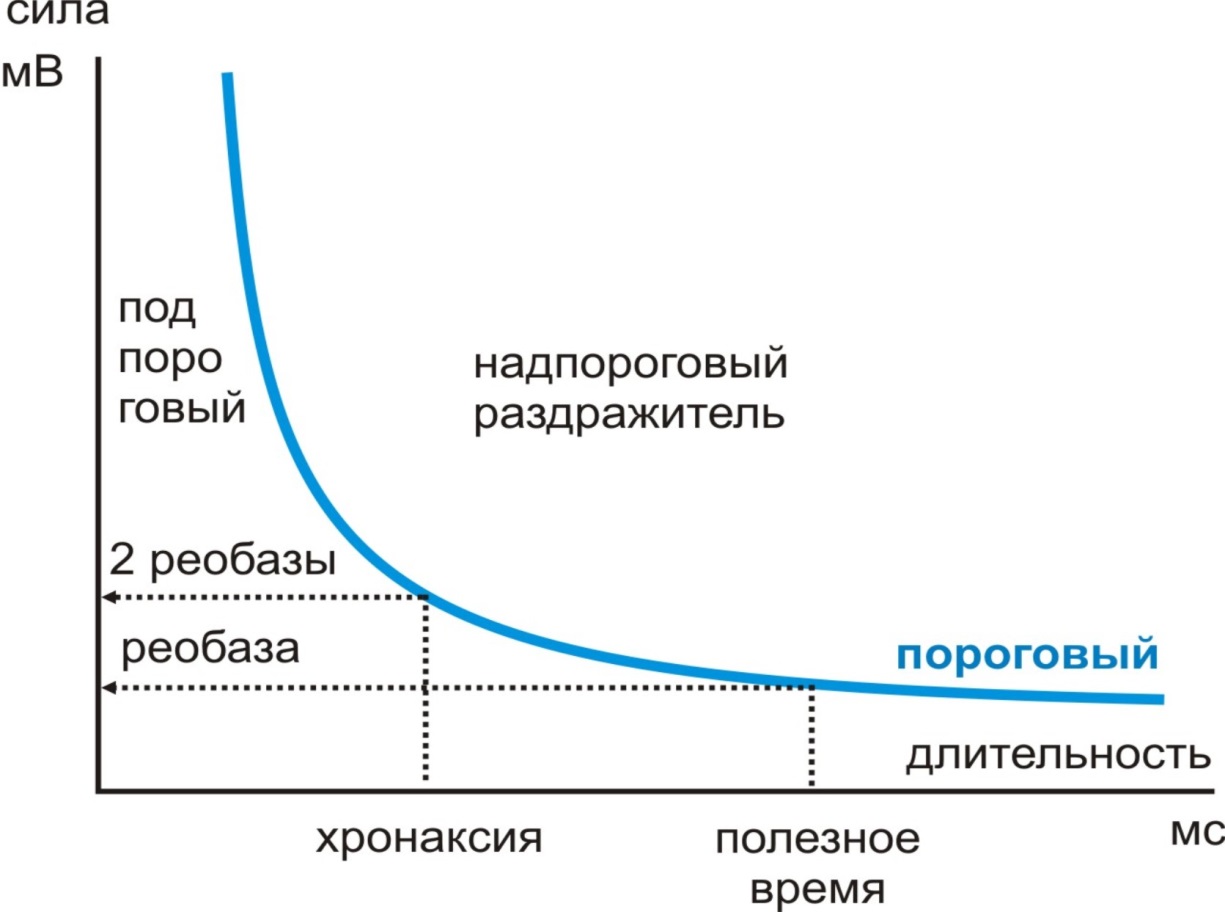
Рис. 7. Кривая «сила-длительность»
Заметьте, что сила раздражителя может быть так мала, что как бы он долго ни действовал, возбуждения не будет. С другой стороны, длительность действия раздражителя может быть такой минимальной, что каким бы он сильным ни был, возбуждение тоже не будет. Поэтому ток высокой частоты оказывает тепловой, а не электрический эффект.
Минимальная сила порогового раздражителя носит название реобазы, а минимальная длительность действия этого раздражителя – полезное время (т.е. время, достаточное для возбуждения). Для оценки функциональной подвижности ткани определяют хроноксию – время действия раздражителя силой в две реобазы (Л. Лапик, 1909). Поскольку возбудимость нервной и мышечной ткани отличны, определение хронаксии используют для диагностики повреждений центральной и периферической нервной системы.
Особенность действия на возбудимые ткани постоянным током выражается законом полярного раздражения (Э. Пфлюгер, 1859): при замыкании цепи постоянного тока пороговой (надпороговой) силы одиночное возбуждение возникает под катодом (отрицательным электродом), а при размыкании – одиночное возбуждение возникает под анодом (положительным электродом).
Почему возбуждение при замыкании цепи постоянного тока возникает под катодом, и почему оно единичное?
Катод несет противоположенный заряд по отношению к положительно заряженной внешней поверхности мембраны клетки. Поэтому под катодом при замыкании цепи постоянного тока пороговой (реобаза) или надпороговой силы возникает деполяризация, и при достижении (КУД) развивается возбуждение. Однако после возбуждения продолжающееся действие катодного тока вызывает инактивацию Na+ каналов: КУД смещается, мембрана становится рефрактерной, несмотря на сохраняющуюся деполяризацию (рис. 8). Тоническое изменение мембранного потенциала под катодом носит название катэлеткротона.
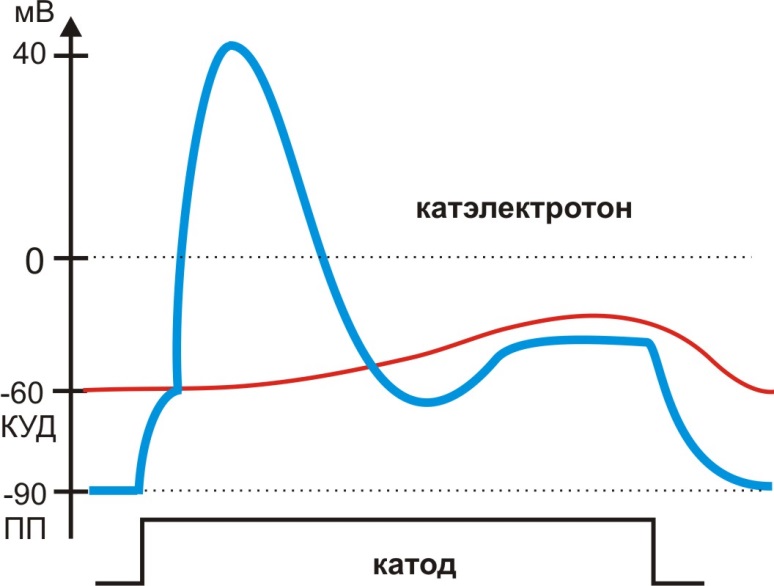
Рис. 8. Изменение мембранного потенциала под катодом, катэлектротон
Развитие рефрактерности возбудимой мембраны под катодом приводит к уменьшению ПД нервов, снижению силы сокращения мышц в ответ на действие надпороговых ритмических раздражителей, что носит название катодической депрессии Вериго (Б.Ф. Вериго, 1883).
При замыкании цепи электрического тока под анодом развивается гиперполяризация - тоническое изменение мембранного потенциала, которое носит название анэлектротона (рис. 9). Под влиянием анодного тока Na+ каналы переходят в состояние экзальтации: КУД, который определяет величину потенциала открытия Na+ каналов, становится больше ПП. Поэтому размыкание цепи постоянного тока возвращает мембранный потенциал к исходной величине ПП, что вызывает открытие потенциалзависимых Na+ каналов – возникает анодно-размыкательное возбуждение (рис. 9).
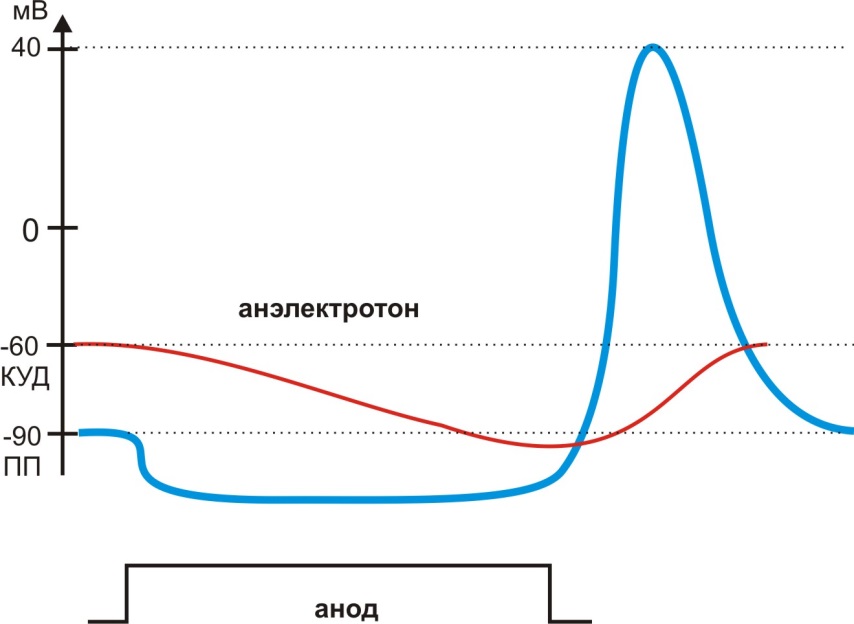
Рис. 9. Изменение мембранного потенциала под анодом, анэлектротон
Потенциал действия способен не только возникать в локальном месте, но и распространяется по мембране клеток возбудимых тканей. Рассмотрим, как протекает этот процесс по мембране нервного волокна.
В немиелинизированных волокнах ионные каналы равномерно распределены по длине нервного отростка. В месте действия порогового раздражителя (ПР) развивается деполяризация и происходит овершут (смена заряда мембраны). Это приводит к возникновению ЭДС (электродвижущих сил), направленных в обе стороны вдоль мембраны. Сила ЭДС такова, что вызывает возбуждение в соседних участках мембраны. Процесс распространяется последовательно по всей мембране нервного волокна (рис. 10).
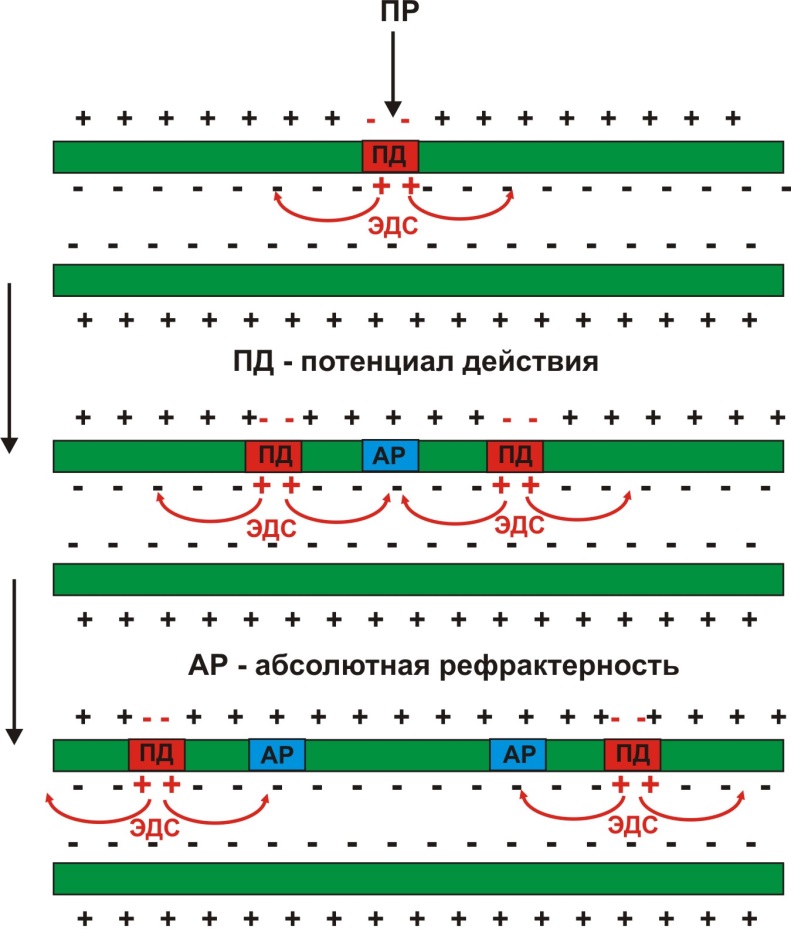
Рис. 10. Распространение возбуждения по немиелинизированному нервному волокну
В целостном организме проведение возбуждения обычно происходит в одном направлении: от сомы нейрона по аксону к нервному окончанию (синапсу).
В миелинизированных нервных волокнах, покрытых шванновскими оболочками (миелином), ионные каналы сконцентрированы в перехватах Ранвье. Это увеличивает амплитуду ПД, ЭДС и, следовательно, на большее расстояние возможна деполяризация соседних участков мембраны. Поэтому миелинизация нервных волокон обеспечивает высокую скорость проведения возбуждения. Возбуждение в миелинизированных нервных волокнах «перепрыгивает» от одного перехвата Ранвье к другому, что получило название сальтаторного проведения возбуждения (рис. 11).
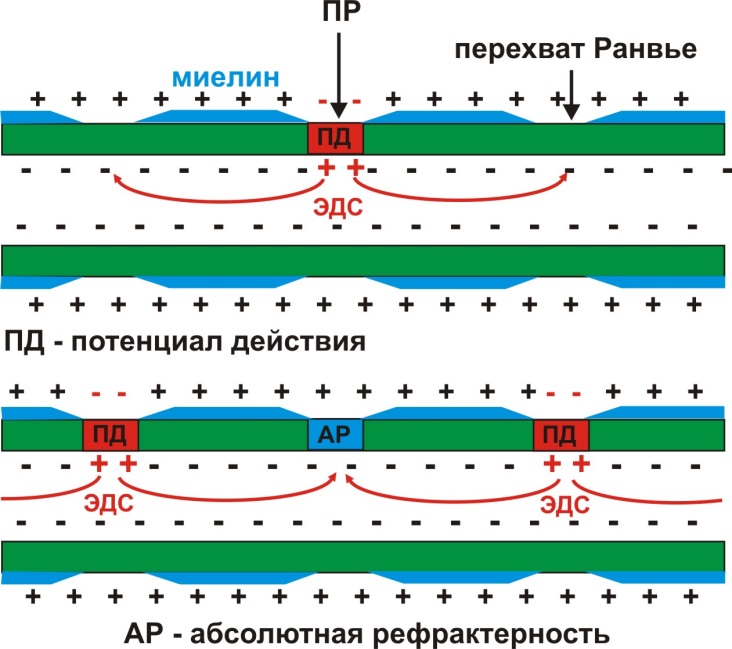
Рис. 11. Сальтаторное проведение возбуждения в миелинизированных нервных волокнах
Демиелинизация нервных волокон в ЦНС приводит к гибели нейронов и развитию рассеянного склероза.
Можно выделить основные закономерности проведения возбуждения по отдельным нервным волокнам:
1. Возбуждение может проводится в обе стороны (обычно в целостном организме возбуждение распространяется векторно: от нервного окончания к соме нейрона, и от сомы по аксону – к нервному окончанию);
2. Амплитуда распространяющегося потенциала действия максимальна, не зависимо от длины нервного волокна (закон «все или ничего»);
3. Для проведения возбуждения необходимы анатомическая и физиологическая целостность проводника (изменение ионного микроокружения, новокаин, лидокаин нарушают проведение).
В ЦНС и на периферии нервные волокна обычно концентрируются в пучки или нервы. В состав спинномозговых нервов входят десятки тысяч нервных волокон. Используя осциллограф, Дж. Эрлангер и Х. Гассер впервые зарегистрировали потенциал действия смешанного нерва. Оказалось, что потенциал действия нерва имеет форму трехгорбой кривой, что объясняется различиями в скорости проведения возбуждения отдельных нервных волокон, входящих в состав нерва (рис. 12).
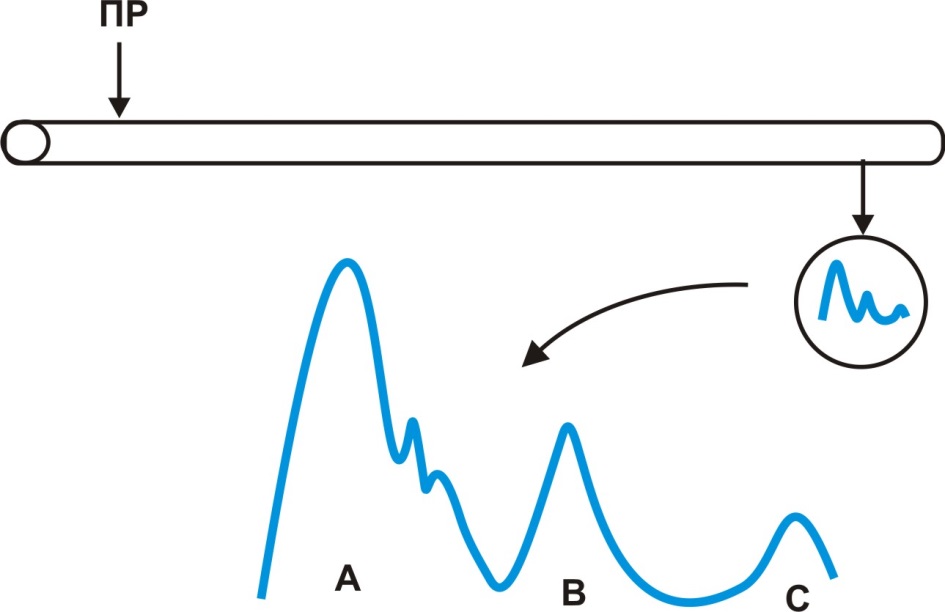
Рис. 12. Потенциал действия нерва
Скорость возбуждения зависит от наличия миелиновой оболочки и диаметра нервного волокна. Исходя из структурно-функциональной особенностей, Дж. Эрлангер и Х. Гассер выделили три типа (в первом типе – несколько подтипов) нервных волокон: Аα - первичные афференты мышечных веретен, двигательные волокна скелетных мышц; Аβ - кожные афференты прикосновения и давления; Аγ - двигательные волокна мышечных веретен; Аδ - кожные афференты температуры и боли; В - симпатические преганглионарные волокна; С- кожные афференты боли, симпатические постганглионарные волокна. Основные характеристики нервных волокон представлены в таблице 3.
Таблица 3
Дата добавления: 2017-03-12; просмотров: 2914;
Поиск по сайту
Узнать еще
- I. Что такое смерть? И что такое жизнь, концом которой является смерть?
- Билет №8 В. 1. Что такое локализация пожара.
- В.1. Что такое боевой участок на пожаре.
- Здоровье, норма – что это такое? Здоровье в гармонии.
- Итак, что такое HTML? И как создать простейшую страницу?
- Какие факторы влияют на выбор конечных параметров пара? Что такое кратность охлаждения в конденсаторе? Каково конечное давление пара на ТЭС и АЭС?
- Каковы основные требования к месту строительства электростанции? Каковы особенности выбора места строительства АЭС? Что такое роза ветров в районе размещения станции?
- Кто или что они такое?

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
