Внизу -стрелолист с надводными, плавающими и подводными листьями
Кролики горностаевой породы имеют белый цвет тела, кроме конца морды, лап, хвоста и ушей. Если выбрить участок белых волос, например, на спине и держать при пониженной температуре (0-1° С), то на выбритом месте отрастает черная шерсть. Если выбрить часть черных волос и поместить кролика в условия повышенной температуры, то вновь отрастает белая шерсть.
Связано это с тем, что для каждого участка тела характерен свой уровень кровообращения и соответствующее варьирование температуры тела, в зависимости от чего формируется или деградирует черный пигмент - меланин (рисунок). Генотип при этом остается одинаковым.

Рисунок - Клубни картофеля, образующиеся над землей при затенении стебля
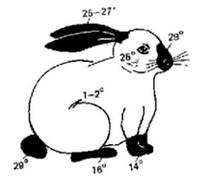
Рисунок - Карта температурных порогов пигментации у кролика
У модификаций описаны следующие свойства:
1. Степень выраженности модификации пропорциональна силе и продолжительности действия на организм вызывающего модификацию фактора. Эта закономерность коренным образом отличает модификации от мутаций, особенно от генных.
2. В подавляющем большинстве случаев модификация представляет собой полезную, приспособительную реакцию организма на тот или иной внешний фактор. Это можно видеть на примере почти всех перечисленных выше и многих других модификаций микробов и человека.
3. Адаптивными бывают только те модификации, которые вызываются обычными изменениями природных условий, множество раз встречавшимися особям данного вида на протяжении его прошлой эволюционной истории. Если же организм попадает в необычные обстоятельства, с которыми его предкам сталкиваться не приходилось, то возникают модификации лишенные приспособительного значения.
4. Не имеют приспособительного значения (а нередко представляют даже настоящие уродства) модификации, вызываемые экстремальными экспериментальными воздействиями, особенно химическими и физическими факторами, с которыми организм не сталкивается в природе. Индуцированные таким образом модификации часто называют морфозами.Если действовать на личинок или куколок дрозофилы рентгеновскими или ультрафиолетовыми лучами, а также предельно переносимой температурой, то у развивающихся мух наблюдаются разнообразные морфозы, характер которых зависит от индуцирующего фактора и его интенсивности, а также от стадии развития организма в момент воздействия. Некоторые из этих морфозов очень похожи на изменения, вызываемые мутациями известных генов. Так, под влиянием теплового шока, которому подвергались предкуколки и куколки, были получены мухи с закрученными кверху крыльями, с вырезками на крыльях, с расставленными крыльями, с крыльями малых размеров, фенотипически неотличимые от мух нескольких мутантных линий дрозофилы (опыты Митчелла). Такие модификации, напоминающие проявление известных генов, получили название фенокопий.
5. Варьирование степени стойкости модификаций. В отличие от высокой константности мутаций, модификации обладают разной степенью стойкости. Многие из них обратимы, т.е. возникшее изменение постепенно исчезает, если устранено вызвавшее его воздействие. Так, загар у человека проходит, когда кожа перестает подвергаться инсоляции, объем мышц уменьшается после прекращения тренировки и т.д.
Лишь очень редко модификация затрагивает, тоже постепенно сходя на нет, ряд поколений, но поколений не половых, а получающихся при вегетативном или партеногенетическом размножении. Такие длительные модификации описаны например, у инфузорий-туфелек. Вначале они выдерживали концентрацию мышьяковистой кислоты не выше 1,1%. Однако, переводя их во все более крепкие растворы, удалось добиться, что они стали переносить даже 5% концентрацию яда. После прекращения воздействия, устойчивость туфелек к мышьяковистой кислоте медленно снижалась, но только через 10 с половиной месяцев она опустилась до исходного уровня, т.е. модификация исчезла лишь приблизительно за 600 вегетативных поколений.
6. Ненаследственный характер модификаций. В отличие от мутаций, модификации не передаются по наследству. Это положение наиболее остро обсуждалось на протяжении всей истории человечества. Полагали, что наследоваться могут любые изменения организма, как врожденные, так и приобретенные в течение жизни. Даже Дарвин признавал возможность наследования некоторых модификационных изменений.
Первый серьезный удар представлению о наследовании приобретенных признаков нанес А. Вейсман.
Иллюстрируя положение о модификациях, А. Вейсман поставил следующий опыт, доказывающий ненаследование приобретенных признаков. На протяжении 22 поколений он отрубал белым мышам хвосты и скрещивал их между собой. В общей сложности было обследовано 1592 особи и ни разу не было обнаружено укорочения хвоста у новорожденных мышат. В подобном эксперименте, результаты которого были опубликованы в 1913 году, в сущности не было необходимости, поскольку события, аналогичные экспериментам Вейсмана часто встречаются и в обычнойжизни людей. "Результаты преднамеренных повреждений у человека, сделанные из ритуальных или "эстетических" соображений – обрезание, протыкание ушей, губ, носовой перегородки, удаление зубов, уродование ступней, черепа и т.д., как известно не наследуются".
В России в 1930-е-1950-е годы получили широкое распространение ошибочные утверждения Лысенко и его последователей о наследовании "приобретенных признаков", т.е. соматических изменений, возникающих в течение жизни особи под влиянием факторов среды, а затем способных адекватно передаваться ее потомкам, т.е. проявляться у них в таком же виде, как у родителя, даже без действия этих факторов.
Множество тщательных опытов, проведенных на разных организмах, показало ненаследуемость модификации, и исследования такого рода представляют теперь лишь исторический интерес.
В наше время Ф. Криком сформулирована т.н. "центральная догма молекулярной биологии", согласно которой перенос информации возможен только от генетического материала к генным продуктам - белкам, но не в обратном направлении.
Норма реакции.Фенотип формируется за счет взаимодействия двух факторов: генотипа и внешней среды.
Свойство данного генотипа обеспечивать в определенных пределах изменчивость онтогенеза в зависимости от меняющихся условий среды, называют нормой реакции. Иначе говоря, амплитуда возможной изменчивости в реализации генотипа выражает норму реакции.
Норму реакции наблюдать лучше всего у организмов с одинаковым генотипом, например у вегетативно размножающихся растений и однояйцевых близнецов. В этом случае можно выявить норму реакции генотипа в наиболее "чистом" виде.
Полностью охарактеризовать норму реакции, присущую тому или иному генотипу, практически невозможно, т.к. для этого пришлось бы изучить как изменяется фенотип особей данного генотипа во всех разнообразных условиях среды, в каких они могут оказаться. Но более частные проявления нормы реакции нередко необходимо знать. В селекции, направленной на создание новых или совершенствование существующих форм полезных человеку организмов, постоянно возникает потребность установить различия в реакции тех или иных сортов возделываемого растения на качество почвы, сроки посева, наличие удобрений.
Какова генетическая обусловленность нормы реакции?
Некоторые из факторов, способные обеспечить варьирование признаков в пределах нормы реакции, можно перечислить:
1. Полигенная детерминация признака и реакции организма.
2. Плейотропность действия гена.
3. Зависимость проявления мутации от условий среды.
4. Гетерозиготность организма, вследствие чего у некоторых генов могут изменяться отношения доминирования.
5. Взаимодействие генов, которое происходит на уровне генных продуктов – субъединиц белковых молекул.
6. Альтернативные пути развития в системе онтогенеза и биосинтезов в клетке. Блокирование одного пути компенсируется другим.
Литература
1. Айала, Ф. Современная генетика / Ф. Айала, Дж. Кайгер. – М.: Мир, 1987. – Т.1. – 295 с; Т.2. – 368 с; Т.3.
2. Алиханян, С. И. Общая генетика / С. И. Алиханян, А. П. Акифьев,
Л. С. Чернин. – М.: Высш. шк., 1985.
3. Бокуть, С. Б. Молекулярная биология: молекулярные механизмы хранениия, воспроизведения и реализации генетической информации / С. Б. Бокуть, Н. В. Герасимович, А. А. Милютин. – Мн.:Высш. шк., 2005.
4. Дубинин, Н. П. Общая генетика / Н. П. Дубинин. – М.: Наука, 1986.
5. Жимулев, И. Ф. Общая и молекулярная генетика / И. Ф. Жимулев. – Новосибирск: Изд-во Новосибирского ун-та, 2002.
6. Жученко, А. А. Генетика / А. А Жученко, Ю. Л. Гужов,
В. А. Пухальский. – М.: Колос, 2004.
Дата добавления: 2017-03-12; просмотров: 2383;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
