Молекулярно-генетические аспекты апоптоза.
Существует несколько путей реализации апоптоза. 1. Важное место занимает путь, опосредованный физиологическими индукторами, действие которых реализуется через клеточные рецепторы, специально предназначенные для включения программы апоптоза. Этот путь передачи сигнала запрограммированной клеточной смерти схематически можно изобразить следующим образом: индукторы ==> рецепторы ==> адаптеры ==> инициирующие каспазы ==> регуляторы ==> эффекторные или казнящие каспазы. Так, рецептор, обозначаемый Fas, взаимодействуя с соответствующим лигандом (лигандом FasL), трансмембранным белком Т-киллера, активируется и запускает программу смерти клетки, инфицированной вирусом. Тем же путем при взаимодействии с лигандом FasL на поверхности ТН-1-лимфоцитов или с антителом к Fas-рецептору погибают ставшие ненужными выздоровевшему организму В лимфоциты, продуценты антител, несущие Fas-рецептор. FasL- лиганд, относящийся к многочисленному семейству фактора некроза опухолей (TNF - tumor necrosis factor). Это семейство гомотримерных лигандов, кроме FasL и TNFa , включает TNFb (лимфотоксин), TRAIL (Apo2L), CD40L, CD27L, CD30L, OX40L. Fas - член семейства рецепторов TNF. Все они представлены трансмембранными белками, которые внеклеточными участками взаимодействуют с тримерами лигандов-индукторов. Взаимодействие рецептора и лиганда приводит к образованию кластеров рецепторных молекул и связыванию их внутриклеточных участков с адаптерами. Адаптер, связавшись с рецептором, вступает во взаимодействие с эффекторами, пока еще неактивными предшественниками протеаз из семейства инициирующих каспаз. Взаимодействие адаптера с рецептором и эффектором осуществляется через гомофильные белок-белковые взаимодействия небольших доменов: DD (death domain - домен смерти), DED (death-effector domain - домен эффектора смерти), CARD (caspase activation and recruitment domain - домен активации и рекрутирования каспазы). Все они имеют сходную структуру, содержат по шесть a-спиральных участков. Домены DD участвуют во взаимодействии рецептора Fas c адаптером FADD (Fas-associated DD-protein) и во взаимодействии рецепторов TNFR1 и DR3 (death receptor 3) с адаптером TRADD (TNFR1-associated DD-protein). Домены DED участвуют во взаимодействии адаптера FADD с прокаспазами 8 и 10. Адаптер RAIDD (RIP-associated Ich-1/CED-3 homologous protein with a death domain, RIP - receptor interacting protein) связывается с прокаспазой-2 через CARD-домены. Наиболее подробно охарактеризована прокаспаза-8 (FLICE/MACH/Mch5), рекрутируемая рецептором Fas через адаптeр FADD. Образуются агрегаты FasL - Fas - FADD - прокаспаза-8. Подобные агрегаты, в которых происходит активация каспаз, называются апоптосомами, апоптозными шаперонами, или сигнальными комплексами, индуцирующими смерть (DISC - death-inducing signaling complex). Каспазы - семейство цистеиновых протеиназ, расщепляющих свои субстраты по остаткам аспартатовой кислоты. Каспазы присутствуют в цитоплазме в виде проэнзимов и активируются до полностью функциональных протеаз путем расщепления проэнзима на большую и малую субединицы и дальнейшего отщепления от них N-концевых доменов. Затем субъединицы собираются в тетрамер с двумя активными центрами. Расщепление прокаспаз могут осуществлять различные протеазы, в том числе и другие каспазы. По выполняемой каспазами функции их можно разделить на две основные группы: инициаторные каспазы (8, 9 и10) и вторая группа - эффекторные каспазы (3, 6 и 7). После того, как каспазы из первой группы активируют эффекторные каспазы, процесс, запущенный программой смерти, оказывается необратимым. Расщепление 3, 6 и 7 каспазами ряда ключевых субстратов, в частности DFF45/ICAD -ингибитора нуклеазы DFF40/CAD (осуществляется каспазой 3), ламинов - ядерных цитоскелетных белков (осуществляется каспазой 6) и т.д., приводит к фрагментации ДНК и деструкции клетки. Существуют другие пути активации каспазы-8 - с участием рецепторов TNFR1 и DR3. Однако эти пути, включаемые одним и тем же адаптером TRADD, конкурируют с параллельными путями активации ядерных факторов транскрипции NF-єB (nuclear factor kappa B) и JNK/AP-1 (JNK, Jun-N-концевая киназа, является компонентом митоген-активируемого киназного пути, ведущего к активации фактора транскрипции AP-1), зависимыми от адаптеров RIP и TRAF (TNFR1-associated factor), под контролем этих факторов транскрипции находится синтез белковых регуляторов, которые блокируют TNF- или Apo3L-индуцированную активацию каспазы-8. Предполагаются следующие пути передачи про- и антиапоптозных сигналов: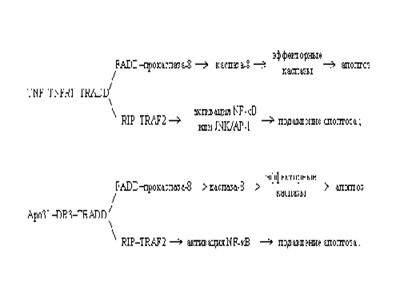 2. Существует путь передачи сигнала апоптоза с участием эндоплазматического ретикулума (ЭР). В ЭР локализована прокаспаза-12. Нарушение внутриклеточного Ca2+-гомеостаза добавкой тапсигаргина или Ca2+-ионофорного антибиотика А23187 ведет к апоптозу клеток, вызванному превращением прокаспазы-12 в каспазу-12. ЭР-зависимый апоптоз связан с болезнью Альцгеймера: кортикальные нейроны мышей, дефицитных по каспазе-12, устойчивы к апоптозу, индуцированному І-амилоидным белком, но не к апоптозу с участием рецепторов плазматической мембраны или митохондриального цитохрома С. 3. Цитотоксические лимфоциты, Т-киллеры, могут вызывать апоптоз у инфицированных клеток с помощью белка перфорина. Полимеризуясь, перфорин образует в цитоплазматической мембране клетки-мишени трансмембранные каналы, по которым внутрь клетки поступают TNFb , гранзимы (фрагментины) - смесь сериновых протеаз. Существенным компонентом этой смеси является гранзим В - протеолитический фермент, превращающий прокаспазу-3 в активную каспазу-3. 4. Высвобождаемый из митохондрий цитохром С вместе с цитоплазматическим фактором APAF-1 (apoptosis protease activating factor-1) участвует в активации каспазы-9. APAF-1 - белок с молекулярной массой 130 кДа, содержащий CARD-домен (caspase activation and recruitment domain) на N-конце и 12 повторяющихся аминокислотных WD-40-последовательностей (WD - дипептид из триптофана и аспартата) на С-конце, образует комплекс с прокаспазой-9 в присутствии цитохрома С и dATP или АТР. К наиболее охарактеризованным WD-белкам относится ?-cубъединица G-белков. Из этих субъединиц собираются жесткие, симметричные структуры, наподобие веера или пропеллера. WD-повторы свойственны белкам, участвующим в регуляции деления и дифференцировки эукариотических клеток, транскрипции генов, модификации мРНК, трансмембранной передачи сигналов, слияния мембранных везикул. Среди прокариот WD-белки обнаружены у цианобактерий. APAF-1 играет роль арматуры, на которой происходит аутокаталитический процессинг каспазы-9. Предполагается, что в результате зависимого от гидролиза dATP (или АТР) конформационного изменения APAF-1 приобретает способность связывать цитохром С. Связав цитохром С, APAF-1 претерпевает дальнейшее конформационное изменение, способствующее его олигомеризации и открывающее доступ CARD-домена APAF-1 для прокаспазы-9, которая тоже содержит CARD-домен. Так образуется конструкция, называемая тоже апоптосомой, с молекулярной массой > 1,3 млн дальтон, в составе которой - не менее 8 субъединиц APAF-1. Благодаря гомофильному CARD-CARD-взаимодействию с APAF-1 в эквимолярном соотношении связывается прокаспаза-9, а затем прокаспаза-9 связывает прокаспазу-3. Пространственное сближение молекул прокаспазы-9 на мультимерной арматуре из APAF-1-цитохром-С-комплексов, по-видимому, приводит к межмолекулярному протеолитическому процессингу прокаспазы-9 с образованием активной каспазы-9. Сходный механизм предложен для активации прокаспазы CED-3 у нематоды Caenorhabditis elegans - аналога прокаспазы-9 млекопитающих. Альтернативный вариант - прокаспаза-9, связавшись с апоптосомой, может принять конформацию, которая приводит к внутримолекулярному процессингу (самоактивации). Зрелая каспаза-9 затем расщепляет и активирует прокаспазу-3. Мутантный APAF-1, лишенный WD-40-повторов, активирует прокаспазу-9, но не способен к рекрутированию и активации прокаспазы-3. Флавопротеин AIF, будучи добавленным к изолированным ядрам из клеток HeLa, вызывает конденсацию хроматина и фрагментацию ДНК, а при добавлении к изолированным митохондриям печени крыс - высвобождение цитохрома С и каспазы-9. Микроинъекция AIF в интактные фибробласты крыс приводит к конденсации хроматина по переферии ядра, разрыву ДНК на крупные фрагменты длиной 50 т.п.н. и больше, снижению мембранного потенциала в митохондриях и переходу фосфатидилсерина из внутреннего слоя цитоплазматической мембраны в наружний. Ни один из этих эффектов AIF не предотвращается пептидным ингибитором каспаз N-бензоилоксикарбонил-Val-Ala-Asp.трифторметилкетоном (Z-VAD.fmk), который предотвращает апоптоз, индуцированный микроинъецированным цитохромом С. Эти данные показывают, что AIF является митохондриальным эффектором апоптоза у животных, действующим независимо от каспаз (Susin S. A. et al, 1999). Кроме рассмотренных компонентов, при нарушении наружной мембраны митохондрий из межмембранного объема выделяется термолабильный фактор, вызывающий необратимое превращение ксантиндегидрогеназы в ксантиноксидазу. Фактор устойчив к ряду испытанных ингибиторов протеаз, включая каспазы, сериновые и металлопротеазы. Ксантиндегидрогеназа катализирует зависимое от NAD+ окисление ксантина до гипоксантина и последующее окисление гипоксантина до мочевой кислоты. Ксантиноксидаза катализирует те же реакции, но не с NAD+, а с О2 в качестве акцептора электронов. При этом образуются О2A, Н2О2, а из них - и другие активные формы кислорода (АФК), которые разрушают митохондрии и являются мощными индукторами апоптоза. Механизмы образования АФК, конечно, не ограничиваются ксантиноксидазной реакцией. Главным источником АФК в клетках являются митохондрии. Резкое увеличение АФК происходит при возрастании мембранного потенциала в митохондриях, когда снижено потребление ATP и скорость дыхания лимитируется ADP. Доля электронного потока через дыхательную цепь митохондрий, идущая на образование О2A, достигает 1-5 %. Цитоплазматическая мембрана макрофагов и нейтрофилов, как уже отмечалось, содержит О2A - генерирующую NADPH-оксидазу. В зависимости от пути, по которому осуществляется активация каспаз, различают несколько типов клеток. Клетки типа I (в частности, линия лимфобластоидных В-клеток SKW и T-клетки линии Н9) подвергаются программированной гибели по пути, зависимому от апоптозных рецепторов плазматической мембраны без участия митохондриальных белков. Клетки типа II (например, линии Т-клеток Jurkat и СЕМ) погибают по пути апоптоза, зависимому от митохондриального цитохрома С. Запрограммированная клеточная смерть, вызванная химиотерапевтическими соединениями, УФ- или ?-облучением, по-видимому, напрямую связана с апоптозной функцией митохондрий: клетки, лишенные генов белка APAF-1 или каспазы-9, устойчивы к химио- и радиационной обработке, но погибают при индукции Fas-рецептора. Некоторые клетки, например, клетки эмбриональной нервной системы, включают механизмы апоптоза, если они испытывают дефицит апоптозподавляющих сигналов (называемых также факторами выживания) от других клеток. Физиологический смысл процесса - в элиминации избыточных нервных клеток, конкурирующих за ограниченное количество факторов выживания. Эпителиальные клетки при отделении от внеклеточного матрикса, вырабатывающего факторы выживания, тоже обречены на гибель. Факторы выживания связываются соответствующими цитоплазматическими рецепторами, активируя синтез подавляющих апоптоз агентов и блокируя стимуляторы апоптоза. Некоторые вещества (например, стероидные гормоны) оказывают дифференцированный эффект на различные типы клеток - предотвращают апоптоз одних типов клеток и индуцируют его у других. 5. Взаимодействие клеток с внеклеточным матриксом осуществляется с помощью интегринов. Интегрины - большое семейство гетеродимерных мембранных белков, которые участвуют в адгезии клеток, связывая внутриклеточный цитоскелет с лигандами внеклеточного матрикса. Нарушение адгезии клеток индуцирует апоптоз. Большинство интегринов специфически взаимодействует с трипептидным RGD (аргинин-глицин-аспартат)-мотивом, входящим в состав белков внеклеточного матрикса. Растворимые низкомолекулярные RGD-содержащие пептиды являются эффективными индукторами апоптоза: проникая в клетки, они активируют латентную каспазу-3. Ряд каспаз, включая каспазу-3, содержит RGD-последовательность вблизи активного центра фермента. В молекуле прокаспазы эта последовательность, вероятно, вовлечена во внутримолекулярное взаимодействие, придающее молекуле профермента такую конформацию, при которой протеазная активность не может проявиться. Предположительно RGD-последовательность взаимодействует с последовательностью DDM (аспартат-аспартат-метионин), локализованной вблизи участка протеолитической активации прокаспазы-3. Низкомолекулярный RGD-пептид, проникая в клетку и вступая в конкурентные взаимоотношения с RGD-последовательностью прокаспазы-3, вытесняет ее из сферы взаимодействия с DDM-последовательностью молекул профермента и индуцирует изменение их конформации, олигомеризацию и аутопроцессинг прокаспазы-3 с образованием активной каспазы-3.
2. Существует путь передачи сигнала апоптоза с участием эндоплазматического ретикулума (ЭР). В ЭР локализована прокаспаза-12. Нарушение внутриклеточного Ca2+-гомеостаза добавкой тапсигаргина или Ca2+-ионофорного антибиотика А23187 ведет к апоптозу клеток, вызванному превращением прокаспазы-12 в каспазу-12. ЭР-зависимый апоптоз связан с болезнью Альцгеймера: кортикальные нейроны мышей, дефицитных по каспазе-12, устойчивы к апоптозу, индуцированному І-амилоидным белком, но не к апоптозу с участием рецепторов плазматической мембраны или митохондриального цитохрома С. 3. Цитотоксические лимфоциты, Т-киллеры, могут вызывать апоптоз у инфицированных клеток с помощью белка перфорина. Полимеризуясь, перфорин образует в цитоплазматической мембране клетки-мишени трансмембранные каналы, по которым внутрь клетки поступают TNFb , гранзимы (фрагментины) - смесь сериновых протеаз. Существенным компонентом этой смеси является гранзим В - протеолитический фермент, превращающий прокаспазу-3 в активную каспазу-3. 4. Высвобождаемый из митохондрий цитохром С вместе с цитоплазматическим фактором APAF-1 (apoptosis protease activating factor-1) участвует в активации каспазы-9. APAF-1 - белок с молекулярной массой 130 кДа, содержащий CARD-домен (caspase activation and recruitment domain) на N-конце и 12 повторяющихся аминокислотных WD-40-последовательностей (WD - дипептид из триптофана и аспартата) на С-конце, образует комплекс с прокаспазой-9 в присутствии цитохрома С и dATP или АТР. К наиболее охарактеризованным WD-белкам относится ?-cубъединица G-белков. Из этих субъединиц собираются жесткие, симметричные структуры, наподобие веера или пропеллера. WD-повторы свойственны белкам, участвующим в регуляции деления и дифференцировки эукариотических клеток, транскрипции генов, модификации мРНК, трансмембранной передачи сигналов, слияния мембранных везикул. Среди прокариот WD-белки обнаружены у цианобактерий. APAF-1 играет роль арматуры, на которой происходит аутокаталитический процессинг каспазы-9. Предполагается, что в результате зависимого от гидролиза dATP (или АТР) конформационного изменения APAF-1 приобретает способность связывать цитохром С. Связав цитохром С, APAF-1 претерпевает дальнейшее конформационное изменение, способствующее его олигомеризации и открывающее доступ CARD-домена APAF-1 для прокаспазы-9, которая тоже содержит CARD-домен. Так образуется конструкция, называемая тоже апоптосомой, с молекулярной массой > 1,3 млн дальтон, в составе которой - не менее 8 субъединиц APAF-1. Благодаря гомофильному CARD-CARD-взаимодействию с APAF-1 в эквимолярном соотношении связывается прокаспаза-9, а затем прокаспаза-9 связывает прокаспазу-3. Пространственное сближение молекул прокаспазы-9 на мультимерной арматуре из APAF-1-цитохром-С-комплексов, по-видимому, приводит к межмолекулярному протеолитическому процессингу прокаспазы-9 с образованием активной каспазы-9. Сходный механизм предложен для активации прокаспазы CED-3 у нематоды Caenorhabditis elegans - аналога прокаспазы-9 млекопитающих. Альтернативный вариант - прокаспаза-9, связавшись с апоптосомой, может принять конформацию, которая приводит к внутримолекулярному процессингу (самоактивации). Зрелая каспаза-9 затем расщепляет и активирует прокаспазу-3. Мутантный APAF-1, лишенный WD-40-повторов, активирует прокаспазу-9, но не способен к рекрутированию и активации прокаспазы-3. Флавопротеин AIF, будучи добавленным к изолированным ядрам из клеток HeLa, вызывает конденсацию хроматина и фрагментацию ДНК, а при добавлении к изолированным митохондриям печени крыс - высвобождение цитохрома С и каспазы-9. Микроинъекция AIF в интактные фибробласты крыс приводит к конденсации хроматина по переферии ядра, разрыву ДНК на крупные фрагменты длиной 50 т.п.н. и больше, снижению мембранного потенциала в митохондриях и переходу фосфатидилсерина из внутреннего слоя цитоплазматической мембраны в наружний. Ни один из этих эффектов AIF не предотвращается пептидным ингибитором каспаз N-бензоилоксикарбонил-Val-Ala-Asp.трифторметилкетоном (Z-VAD.fmk), который предотвращает апоптоз, индуцированный микроинъецированным цитохромом С. Эти данные показывают, что AIF является митохондриальным эффектором апоптоза у животных, действующим независимо от каспаз (Susin S. A. et al, 1999). Кроме рассмотренных компонентов, при нарушении наружной мембраны митохондрий из межмембранного объема выделяется термолабильный фактор, вызывающий необратимое превращение ксантиндегидрогеназы в ксантиноксидазу. Фактор устойчив к ряду испытанных ингибиторов протеаз, включая каспазы, сериновые и металлопротеазы. Ксантиндегидрогеназа катализирует зависимое от NAD+ окисление ксантина до гипоксантина и последующее окисление гипоксантина до мочевой кислоты. Ксантиноксидаза катализирует те же реакции, но не с NAD+, а с О2 в качестве акцептора электронов. При этом образуются О2A, Н2О2, а из них - и другие активные формы кислорода (АФК), которые разрушают митохондрии и являются мощными индукторами апоптоза. Механизмы образования АФК, конечно, не ограничиваются ксантиноксидазной реакцией. Главным источником АФК в клетках являются митохондрии. Резкое увеличение АФК происходит при возрастании мембранного потенциала в митохондриях, когда снижено потребление ATP и скорость дыхания лимитируется ADP. Доля электронного потока через дыхательную цепь митохондрий, идущая на образование О2A, достигает 1-5 %. Цитоплазматическая мембрана макрофагов и нейтрофилов, как уже отмечалось, содержит О2A - генерирующую NADPH-оксидазу. В зависимости от пути, по которому осуществляется активация каспаз, различают несколько типов клеток. Клетки типа I (в частности, линия лимфобластоидных В-клеток SKW и T-клетки линии Н9) подвергаются программированной гибели по пути, зависимому от апоптозных рецепторов плазматической мембраны без участия митохондриальных белков. Клетки типа II (например, линии Т-клеток Jurkat и СЕМ) погибают по пути апоптоза, зависимому от митохондриального цитохрома С. Запрограммированная клеточная смерть, вызванная химиотерапевтическими соединениями, УФ- или ?-облучением, по-видимому, напрямую связана с апоптозной функцией митохондрий: клетки, лишенные генов белка APAF-1 или каспазы-9, устойчивы к химио- и радиационной обработке, но погибают при индукции Fas-рецептора. Некоторые клетки, например, клетки эмбриональной нервной системы, включают механизмы апоптоза, если они испытывают дефицит апоптозподавляющих сигналов (называемых также факторами выживания) от других клеток. Физиологический смысл процесса - в элиминации избыточных нервных клеток, конкурирующих за ограниченное количество факторов выживания. Эпителиальные клетки при отделении от внеклеточного матрикса, вырабатывающего факторы выживания, тоже обречены на гибель. Факторы выживания связываются соответствующими цитоплазматическими рецепторами, активируя синтез подавляющих апоптоз агентов и блокируя стимуляторы апоптоза. Некоторые вещества (например, стероидные гормоны) оказывают дифференцированный эффект на различные типы клеток - предотвращают апоптоз одних типов клеток и индуцируют его у других. 5. Взаимодействие клеток с внеклеточным матриксом осуществляется с помощью интегринов. Интегрины - большое семейство гетеродимерных мембранных белков, которые участвуют в адгезии клеток, связывая внутриклеточный цитоскелет с лигандами внеклеточного матрикса. Нарушение адгезии клеток индуцирует апоптоз. Большинство интегринов специфически взаимодействует с трипептидным RGD (аргинин-глицин-аспартат)-мотивом, входящим в состав белков внеклеточного матрикса. Растворимые низкомолекулярные RGD-содержащие пептиды являются эффективными индукторами апоптоза: проникая в клетки, они активируют латентную каспазу-3. Ряд каспаз, включая каспазу-3, содержит RGD-последовательность вблизи активного центра фермента. В молекуле прокаспазы эта последовательность, вероятно, вовлечена во внутримолекулярное взаимодействие, придающее молекуле профермента такую конформацию, при которой протеазная активность не может проявиться. Предположительно RGD-последовательность взаимодействует с последовательностью DDM (аспартат-аспартат-метионин), локализованной вблизи участка протеолитической активации прокаспазы-3. Низкомолекулярный RGD-пептид, проникая в клетку и вступая в конкурентные взаимоотношения с RGD-последовательностью прокаспазы-3, вытесняет ее из сферы взаимодействия с DDM-последовательностью молекул профермента и индуцирует изменение их конформации, олигомеризацию и аутопроцессинг прокаспазы-3 с образованием активной каспазы-3.
Дата добавления: 2019-09-30; просмотров: 727;
Поиск по сайту
Узнать еще
- Агроэкологические аспекты взаимодействия ходовой части тракторов и автомобилей с почвой
- АНАТОМО-ФИЗИЗИОЛОГИЧЕСКИЕ АСПЕКТЫ ПОТРЕБНОСТИ ВЫДЕЛЯТЬ
- Анатомо-физиологические аспекты занимающихся различного возраста и квалификации
- Анатомо-физиологические аспекты потребностей человека. Человек как предмет изучения анатомии и физиологии
- Аспекты в соционике
- Аспекты использования стоимости капитала
- Аспекты качества продукции
- Аспекты охраны природы

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
