Переноса электронов. (ЭПЦ).
Существуют различные теории биологического окисления:
Перекисная теория биологического оксисления Баха
Окисление глюкозы – Палладин.
Биологическое окисление, или тканевое дыхание – это окислительно-восстановительный процесс разложения субстратов (S), связанный с образованием воды при переносе электронов от Н+ к О2 и выделением Е.
Механизм тканевого дыхания был раскрыт благодаря исследованиям ученых различных стран: Варбург (Германия), Палладин, Бах, (Россия) и др. (конец 19-начало 20 вв.)
Тканевое дыхание – это полиферментная цепь переноса ℮‾ и Н+ от S (донора водорода) на кислород. Эта цепь переносчиков называется дыхательной цепью.
Водород используется организмом как главное топливо для образования энергии. В митохондриях поток электронов от водорода устремляется к конечному акцептору кислороду, при этом образуется молекула воды, которая является конечным продуктом тканевого дыхания. ═> Тканевое дыхание - окисление субстрата, приводящее к его распаду и образованию энергии.
Например, распад ГЛК до конечных продуктов обмена суммарно выражается следующим уравнением: С6Н12О6 + 6О2 ═>6 СО2+6 Н2О + 2850 кдж/моль.
Это превращение осуществляется (как и окисление др. S). путем дегидрирования или отщепления от окисляемого в-ва Н+ и ℮‾ . При каждом дегидрировании Н2 присоединяется к КоЕ (к коферменту ДГ) и КоЕ восстанавливается:
S•Н2+КоЕ"S + КоЕ•Н2
Чаще всего такие реакции происходят в митохондриях. Восстановленные коферменты поступают в дыхательную цепь, где они вновь окисляются.
Напр. НАД•Н2 ═>НАД + 2Н+
Отщепившийся Н+ передается не менее чем через 5 переносчиков (до 80) к концу цепи, где соединятся с молекулой О2 и образует воду. Переход Н+ по ЭПЦ состоит из ряда окислительно-восстановительных реакций. В некоторых из этих реакций образуется достаточно Е для образования АТФ.
Сопряжение тканевого дыхания (образование Н2О) и фосфорилирования (образования АТФ) называетсяокислительным фосфорилированием.
Структура и функции дыхательной цепи (ЭПЦ).
Дыхательная цепь состоит из следующих переносчиков : р+ и е‾
1) Пиридинзависимые ДГ,
их коферментом служат НАД или НАДФ, производные никотиновой кислоты (вит РР) и аденина. Их > 150.
S•H2 + НАД D S + НАД • Н2
Субстрат (НАДФ) окисл. (НАДф•Н2)
восст. форма.
Присоединение и отдача е‾ и Н+ происходит за счет амида никотиновой кислоты. НАД главным образом находится в митохондриях, используется для переноса ℮‾ и р+ от S к О2; НАДФН2 используется главным образом при синтезе Ж.К.
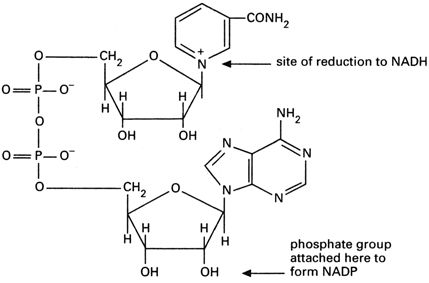 + Н3РО4 = НАДФ
+ Н3РО4 = НАДФ
НАДФ никотинамидадениндинуклеотид никотинамидадениндинуклеотидфосфат
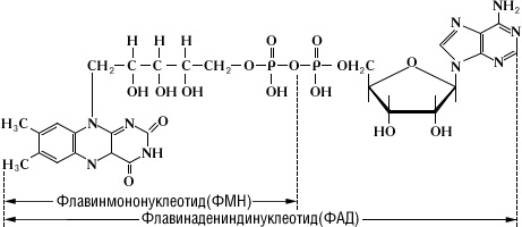
2) Флавиновые ферменты ≈ 30
Простетической группой является ФАД и ФМН. Это следующие после пиридинзависимых ДГ переносчики
НАД ∙Н2 + ФАД (ФМН) ↔ НАД +Фн∙Н2
Фн 2Н+ 2℮‾
ФАД играет роль первичного переносчика р+ и е+ (окисление сукцината ЖК) ФАД, ФМН в отличи от НАД прочно связывается с субстратом
3) Убихинон, КоQ10, Q переносчик р+ и ℮‾ от Фн
Фн ∙Н2 + Q ↔ Фн + Q ∙Н2
2Н 2℮‾ убихинол
4) Цитохромы переносят только электроны, имеют простетическую группу подобную гемму гемоглобина, отличаются белковым компонентом и некоторыми радикалами в простетической группе. Включается в такой последовательности:
cytb¦cytс1¦cytc¦cyt aa3
b1 c1 c – это промежуточные переносчики е.
аа3 (цитохромоксидаза) – конечный переносчик, называется терминальным дыхательным ферментом, т.к. непосредственно взаимодействует с О2 и содержит ионы Сu+1 → Сu+2 , основную функцию выполняет гемовое железо
Fe+3 (aa3) +е (c) D Fe+2 (aa3)
2 Fе+2 + ½ О2о D 2 Fе+3 + О-2
О-2 +2Н+ D Н2О
На участках между НАД и Q происходит перенос 2℮‾, а с Q на суtb – только 1℮‾, поэтому в цепи на 1 НАД участвуют 2 ряда суt.
Упрощенная схема ЭПЦ.
S•H2 (НАД – завис.) S•H2 (ФАД - завис.)
$ $℮‾
НАД •H2 ¦Фн•H2 ¦Q•H2 ¦суtb¦суtc1¦суtc ¦суtaa3 + 1/2 О2 "Н2О
9 ------------------------------------------------¦ 2Н+
Н2 от первичных доноров (S) вводится в дыхательную цепь с участием ФАД- и НАД- зависимых ДГ.
НАД- зависимые ДГ переносят Н+ с НАДН2 на Фп, затем на убихинон, ФАД-зависимые ДГ переносят Н+ сразу на убихинон.
На стадии образования убихинона сливаются оба потока Н+ вводимых в дыхательные цепь НАД- и ФАД- зависимыми ДГ.
Затем пути р+и е‾ расходятся: р поступают в матрикс, ℮‾ через цитохромы передаются на О2. В результате тканевого дыхания в сутки образуется ~ 300-400 мл. воды (так называемая «метаболическая» вода).
Строение митохондрий.
Обычно митохондрии имеют форму цилиндра с закругленными краями; дл. 1-4 мкм; d=0,3 – 0,7 мкм. В различных клетках форма, размеры, число митохондрий различно. Митохондрии имеют наружную и внутреннюю мембраны по типу мешок в мешке. Внутренняя мембрана (ВМ) имеет выпячивания кристы. Содержимое пространства, ограниченное вн.м., называется матриксом.
Наружная мембрана свободно проницаема для молекул с Мr до 5000Да. Проницаемость ВМ ограничена и избирательна и определяется наличием специальных транспортных систем. НАД-зависимые и некоторые ФАД-зависимые ДГ встроены во ВМ митахондрию, их АЦФ обращены внутрь матрикса, т.к. именно в матриксе находятся их S.
Сопряжение тканевого дыхания и фосфорисуtлирования.
Для образования АТФ необходима затрата энергии. В ЭПЦ она образуется на 3-х участках
↓ ↓ ↓
НАД•H2¦Фп¦Q•H2¦суtb¦суtc1¦суtc¦суtaa3¦H2o
Т.о. окислительное фосфорилирование – это синтез АТФ.
Ф-ты ЭПЦ фиксированы во ВМ митохондрий т.о., что их действие векторное (направленное).
Основным проявлением векторности в ЭПЦ является перенос протонов Н+ с внутренней стороны мембраны на наружную.
С НАД•Н2 ℮‾ переходят на Фп, Н+ освобождаются в ММП;
℮‾ с Фн•H2 переходят на Q, а протоны опять поступают в ММП
Q•Н2 (убихинол) также высвобождает Н+ в ММП, электроны переходят на цитохромы.
Фп и Q получают протоны из матрикса, а НАД•H2 от S.
Т.о., при переходе всего 2х ℮‾ в ММП переходят 6Н+. Цепь переноса е‾ работает как протонный насос, перекачивая Н+ из матрикса в ММП; В результате по сторонам мембраны создается разность потенциалов со знаком + на НМ.
Электрохимический потенциал заставляет Н+ двигаться в обратном направлении, т.е. с наружной на внутреннюю мембрану. Однако ВМ не проницаема для Н+, за исключением специальных участков, называемых протонными каналами. В них на внутренней стороне располагается фермент Н+-АТФ-синтетаза, катализирующий реакцию: АДФ+Ф=АТФ+Н2О – образующаяся АТФ при помощи фермента транслоказы транспортируется из матрикса на наружную сторону мембраны и попадает в цитозоль. Транслоказа одновременно переносит АТФ в обратном направлении (из цитозоля в матрикс).
Все эти данные составляют основные положения хемиоосмотической теории Митчелла (1961г.) – теории окислительного фосфорилирования.
Каждая пара протонов способствует образованию 1 мол. АТФ.
Т.о, из НАД•H2¦6Н+¦3 АТФ,
Фп•Н2 ¦ 4Н+¦2 АТФ.
Разобщающие агенты.
Разобщение процессов тканевого дыхания и фосфорилирования наблюдается при наличии в клетках «протонофоров» - т.е. молекул, способных переносить Н+ через мембрану (ВМ). В этом случае выравнивается градиент концентрации, исчезает движущая сила фосфорилирования. К числу протонов относятся 2,4 динитрофенол, гормоны щитовидной железы, жирные кислоты и др.
Соединения, разобращающие окисление и фосфорилирования, называются разобщителями.
В митохондриях дыхание не всегда сопряжено с фосфорилированием. Такой путь окисления S-в в дыхательные цепи Ленинджером был назван свободным окислением или нефосфорилирующим окислением. В противном случае ЭПЦ работает «вхолостую», вся энергия окисляемых S переходят в тепловую. Митохондрии становятся своеобразной клеточной «печкой». Это необходимо в тех случаях, когда потребность в теплоте для организма важнее, чем потребность в энергии (при переохлаждении).
Для мышечных клеток производство теплоты – это не основная функция. В организме имеется ткань – «бурый жир», митохондрии, которые специализированы на выработке теплоты. Бурого жира много у новорожденных, с возрастом его количество убывает. Необычная для жира окраска обеспечивается большим содержанием в его клетках митохондрий, в них в 10 раз больше ферментов дыхания, чем ферментов фосфорилирования, т.е. они меньше настроены на производство АТФ, на тканевое дыхание. Разобщение дыхания и фосфорилирования в митохондриях бурого жира ведет к образованию теплоты, согревающей протекающую кровь.
Основные компоненты пищи.
С точки зрения законов термодинамики живые организмы – это термодинамически открытые системы, т.е. такие системы, которые постоянно обмениваются с внешней средой веществами и энергией (закрытые – только энергией). Обмен веществ живых организмов включает:
а) поступление веществ из среды в организм (в результате питания и дыхания).
б) превращение веществ в организме (промежуточный обмен или метаболизма).
в) выделение конечных продуктов обмена.
Масса тела взрослого человека сохраняется постоянной, т.е. масса потребляемых организмом веществ = массе выделяемых из организма веществ за то же время.
В каждой клетке организма происходит постоянный распад ее собственных структурно-функциональных компонентов, за счет чего образуется мономеры: АК, моносахара, ЖК, нуклеотиды и др. Они смешиваются с мономерами, образующимися при распаде продуктов питания, и образуют общий фонд метаболитов организма. Он расходуется по 2-м направлениям:
а) происходит возобновление распавшихся структурно – функциональных компонентов клетки;
б) превращение мономеров в конечные продукты обмена веществ, которые выводятся из организма.
При этом распаде у взрослого человека в сутки выделяется приблизительно 8-12 тысяч кдж. (2000-3000 ккал) энергии. Эта энергия расходуется:
а) на поддержания постоянной температуры тела;
б) на совершение разного рода работы.
Главную долю органических веществ пищи составляет Ж, Б, У, а также минорные компоненты пищи, требующиеся в малых количествах: витамины, минеральные вещества, вода.
Питание – составная часть обмена веществ
У здоровых У больных
 Гигиена питания диетология
Гигиена питания диетология
Среднесуточный рацион взрослого человека ≈ 3000ккал (1ккал=4,19 кДж)
Существует понятие сбалансированное рациональное питание, т.е.соотношение Б:Ж:У: 1:1:4
Среднесуточное потребление энергии с основными пищевыми веществами
у взрослого человека
| Б | 80г | 328 ккал | 1360 кДж |
| Ж | 100г | 930 ккал | 3900 кДж |
| У | 400г | 1640 ккал | 6800 кДж |
| ∑ | 580г | 2898 ккал | 12060 кДЖ |
АК: три 1г, глу 6г, остальные 2-6г.
В зависимости от пола, возраста, климатических условий, профессии эти показатели могут меняться.
Незаменимые компоненты пищи:
Многие пищевые вещества при их дефиците в пище могут синтезироваться клетками организма: жиры из углеводов, углеводы из аминокислот. Такие вещества называются заменимыми.
В ходе эволюции, когда пища содержала готовые витамины и АК, у человеческого организма постепенно отпала необходимость образовывать собственные ферменты для их синтеза, в результате были утрачены соответствующие гены. При этом стали проще метаболические пути, но вместе с тем возникала зависимость организма от поступления некоторых веществ из внешней среды.
Т.о., имеются группы веществ, являющихся для организма незаменимыми. Наличие их в организме жизненно необходимо, синтезировать их организм не в состоянии, поэтому они должны поступать с пищей:
а) некоторые АК
б) некоторые ЖК ( арахидоновая, линолевая, линоленовая)
в) витамины
г) минеральные вещества
д) вода
Аминокислоты делят на 4 групы:
1) незаменимые: вал, лиз, иле, тре, мет, фен, три.
2) частично заменимые: гис, арг,
3) условно заменимые: цис, тир.
4) заменимые: ала, асп, асн, глу, глн, про, гли, сер.
Если в клетке отсутствуют заменимые АК, организм синтезирует ее из других веществ .
Если же в организме отсутствует хотя бы 1 незаменимая АК, то полностью прекращается синтез белков (!)
Частично заменимые АК: Гис и арг синтезируется в организме, однако скорость их образования недостаточна по сравнению с потребностью в них
Условно заменимые АК: могут синтезироваться из незаменимых: тир из фен, цис из мет. Т.о. они заменимы, если в организм в достаточном количестве поступили АК фен и мет.
Незаменимые ЖК: большинство Ж.К. при дефиците их в пище могут синтезироваться из углеводов.
Незаменимыми являются некоторые полиненасыщенные Ж.К.:
Линолевая СН3 – (СН2)4-СН=СН-СН2 СН=СН-(СН2)7-СООН (С18)
Арахидоновая – (С20)
Линоленовая – (С18)
Линолевая и линоленовая Ж.К. являются предшественниками арахидоновой, а из последней синтезируются гормоноподобные вещества – простагландины – высокоактивные биологические вещества.
Минеральные вещества– это хлориды, фосфаты, карбонаты Na+, К+, Са+2, Мg+2, а также микроэлементы Cr, Zn, Со, Сu, Мо и т.д. – поступают с водой и растительной пищей. Их недостаточность возникает редко (железодефицит, анемия, эндемичный зоб)
Вода – небольшое количество воды (≈400г. метаболической воды) образуется в организме при обмене Ж, Б, У, суточная потребность 1750-2200 мл, она поступает в организм с продуктами биологического и небиологического происхождения.
ЛЕКЦИЯ 10
Дата добавления: 2016-11-29; просмотров: 1858;
Поиск по сайту
Узнать еще
- I.1.9 ЯВЛЕНИЯ ПЕРЕНОСА В ГАЗАХ
- Аналогия явления переноса субстанций. Законы молекулярного переноса
- Геодезическая подготовка проекта для переноса его на местность
- Зависимость массовой скорости горения от тепло-и массопереноса (уравнение Зельдовича)
- Законы переноса и принцип движущей силы
- Лекция 3. Явления переноса
- Метод переноса (ПАФОС)
- Методики ускорения переноса умений и навыков в условия соревновательного поединка при подготовке спортсменов высокой квалификации в вашем виде восточных боевых искусств.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
