КЛЕТОЧНАЯ СТЕНКА (КС)
Строение. КС— важный и обязательный структурный элемент подавляющего большинства бактериальных клеток (кроме микоплазм).
КС покрывает всю поверхность бактериальной клетки. Она располагается под капсулой или слизистым чехлом, у бескапсульных клеток — непосредственно контактирует с окружающей средой (рис.14).

Рис. 14. Схема взаиморасположения внешних слоев клетки бактерий:
1 — ЦПМ, 2 — КС, 3 — микрокапсула, 4 — капсула, 5 — слизистый слой.
На долю КС приходится 5–50 % сухого вещества клетки, количество материала КС увеличивается с возрастом.
Пептидогликан (муреин, мукопептид, ПГ) образует опорный скелет бактериальной клетки, составляет основу КС и специфичен только для бактерий. ПГ имеет структуру молекулярной сети, благодаря двум типам связей — гликозидным и пептидным.
Цепочки ПГ образованы чередующимися остатками N–ацетилглюкозамина и N–ацетилмурамовой кислоты, соединенными между собой β–1,4–гликозидными связями (рис. 15).
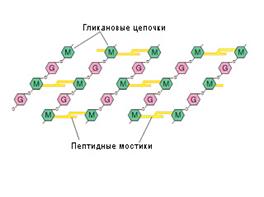
Рис. 15. Гликозидные связи соединяют
N–ацетилмурамовую кислоту (M ) и
N–ацетилглюкозамин G)
в молекуле ПГ
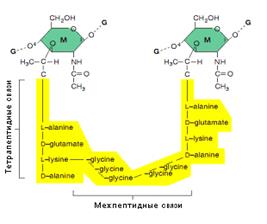
Рис.16. Два набора пептидных цепочек в молекуле ПГ
В молекуле ПГ есть два набора пептидных цепочек — боковые и поперечные.
К N–ацетилмурамовой кислоте присоединен короткий пептидный хвост из 4 аминокислот (тетрапептид). Тетрапептид состоит из чередующихся L– и D– аминокислот (рис. 17). Аминокислоты, участвующие в образовании пептидных связей, варьируют у разных видов бактерий.
Принципиальное значение для пространственной организации ПГ имеет высокое содержание аминокислот с двумя аминогруппами. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей, причем вторые аминогруппы — в формировании межпептидных связей между гетерополимерными цепочками.

Рис. 17. Структура повторяющейся единицы ПГ КС.
Цифры в кружках обозначают:
1, 2 — места полимеризации гликанового остова молекулы;
3 —место присоединения с помощью фосфодиэфирной связи молекулы ТК в КС Грам+ бактерий;
4, 5 —места связывания гликановых цепей с помощью пептидных связей;
6 —место ковалентного связывания (пептидная связь) с липопротеином НМ у Грам- бактерий;
7 — место действия лизоцима
В большинстве случаев в образовании межпептидной связи участвует карбоксильная группа D–аланина одного тетрапептида и свободная аминогруппа диаминокислоты другого (рис.18А). Иногда связь между тетрапептидами разных гликановых цепей осуществляется с помощью других аминокислот (напр., глицина у S. aureus) (рис.18Б).
 Рис. 18. Межпептидные мостики между гетерополимерными цепочками:
Рис. 18. Межпептидные мостики между гетерополимерными цепочками:
Г — N–ацетилглюкозамин;
М — N-ацетилмурамовая кислота;
ала — аланин; глу — глутаминовая кислота; лиз — лизин; мезо–ДАП–мезодиаминопимелиновая кислота; гли — глицин.
Стрелками обозначено место действия пенициллина
Частота «сшивок» гетерополимерных цепей различна, поскольку не все пептидные хвосты участвуют в формировании межцепочечных связей. Часть пептидных хвостов находится в свободном состоянии.
В КС бактерий содержатся структуры и вещества, которых нет у человека, животных и растений:
– N–ацетилглюкозамин;
– N–ацетилмурамовая кислота;
– мезо–диаминопимиелиновая кислота;
– D–аланин;
– D–глутаминовая кислота.
Это «ахилесова пята» бактерий, используемая врачами в борьбе с инфекцией, так как некоторые лекарственные препараты действуют только на КС бактерий, и не затрагивают эукариотических клеток высших организмов.
На ПГ откладываются и его инкрустируют различные вещества. По строению ПГ и по содержанию других веществ в КС Грам+ бактерии отличаются от Грам-. Химический состав и строение КС постоянны для определенного вида и являются важным признаком. В зависимости от строения КС бактерии делятся на две большие группы: грамположительные (Грам+) и грамотрицательные (Грам-) (рис.19).
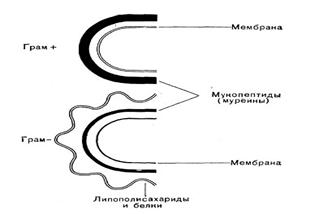
Рис. 19. Схема строения КС
у Грам+ и Грам- бактерий
Особенности КС Грам+ бактерий:
1) мощная и толстая, в зависимости от вида бактерий толщиной 20–60 нм (в 2–3 раза толще, чем у Грам- бактерий);
2) основную часть массы КС составляет ПГ (40–90%);
3) ПГ многослойный(10 слоев);
4) у Грам+ бактерий обнаружено более 100 различных химических типов ПГ. Большинство различий относится к структуре тетрапептида. В образовании боковых пептидных связей у Грам+ бактерий участвует LL–диаминопимиелиновая кислота или лизин;
5) ПГ ковалентно связан с тейхоевыми кислотами (ТК) (от греч. teichos — стенка). ТК — полимерные цепи, состоящие их 8–50 остатков рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки соединены между собой фосфодиэфирными связями. Длинные линейные молекулы ТК могут пронизывать весь ПГ слой, достигая внешней поверхности КС. В этом случае они являются основными антигенами Грам+ бактерий. Свободные гидроксилы фосфорной кислоты придают ТК свойства полианиона и определяют поверхностный заряд клетки. Углеводные компоненты ТК входят в состав клеточных рецепторов для бактериофагов. Липотейхоевые кислоты фиксированы в мембране липофильными концами;
6) нет ЛПС, содержится небольшое количество полисахаридов, липидов и белков; полисахариды и липиды ковалентно связываются с макромолекулами КС; белки формируют на внешней поверхности КС отдельный слой;
7) КС плотно прилегает к ЦПМ, нет периплазматического пространства;
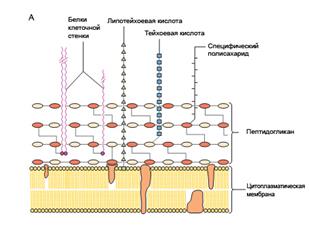
| Б |
|
Рис. 20. Строение КС:
А – у Грам+ бактерий, Б – у Грам- бактерий
8) Грам+ бактерии чувствительны к пенициллину и лизоциму. Пенициллин разрушает тетрапепдидные связи. Лизоцим разрушает гликозидные связи.
Таким образом, основными компонентами КС Грам+ бактерий являются три типа макромолекул: пептидогликаны, тейхоевые кислоты и полисахариды, которые с помощью ковалентных связей образуют сложную структуру с упорядоченной пространственной организацией (рис. 20А).
Особенности КС Грам- бактерий:
1) значительно (в 2–3 раза) тоньше, чем у Грам+; ее толщина 10–20 нм;
2) содержание ПГ значительно меньше, чем у Грам+ бактерий, и составляет в зависимости от вида бактерий 5–10 % сухой массы КС;
3) ПГ однослойный или двуслойный, толщиной 2–3 нм;
4) у всех Грам- бактерий строение ПГ одинаково. В образовании боковых пептидных связей у Грам+ бактерий участвует только мезо–диаминопимиелиновая кислота (лизин отсутствует). Поэтому гетерополимерные цепи между собой связаны редкими поперечными связями через два однотипных тетрапептида.В образовании межпептидной связи участвует мезо–диаминопимиелиновая кислота и D–аланин;
5) ПГ не содержит ТК;
6) ПГ неплотно прилегает к ЦПМ. Только у Грам- бактерий между ЦПМ и ПГ КС есть периплазматическое пространство. Тонкий ПГ соединен белками с наружной мембраной;
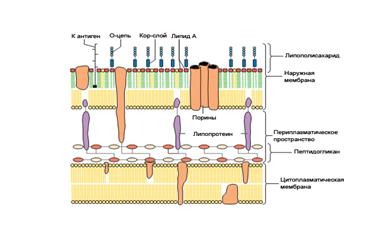
7) КС многослойная, сверху ПГ только у Грам- бактерий находится наружная мембрана (НМ) толщиной 8–10 нм. Она оставляет до 80% сухой массы КС. НМ по строению сходна с внутренней ЦПМ и состоит из липопротеина (ЛП), липополисахарида (ЛПС), фосфолипидов и белков.
Основной компонент НМ — билипидный слой: внутренний слой образован ЛП, а наружный — ЛПС. ЛП, ЛПС и другие липиды связаны ковалентно, ЛП ориентированы липофильными концами наружу.
ЛПС занимает 30–40 % поверхности НМ и состоит из трех компонентов:
– липида А, который «заякоривает» ЛПС в НМ, он содержит глюкозамин и жирные кислоты, придает токсичность липополисахариду и является одним из основных факторов патогенности. Токсические свойства проявляются преимущественно при разрушении бактериальных клеток;
– кор–слоя (лат. core — ядро), одинакового для всех Грам- бактерий, наиболее постоянной частью которого является кетодезоксиоктоновая кислота;
– О–специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями. Она определяет антигенную специфичность, т. е. серовар определенного штамма бактерий, является О–антигеном. Напр., в серологических реакцих было идентифицировано более 1000 сероваров сальмонелл. Определение сероваров бактерий имеет большое значение в бактериологической диагностике и в расшифровке эпидемий (позволяет установить источник инфекции). Полисахаридные цепи дают бактериям преимущества в процессе отбора. Огромное разнообразие полисахаридных цепей объясняется отбором новых мутантных типов О–антигенов. Новые типы дают преимущество, так как хозяин не может обладать антителами против всех антигенов одновременно.
Много различных белков локализовано в НМ Грам- бактерий. Белки НМ делят на основные и минорные. Основные белки (почти 80 % всех белков НМ) представлены небольшим числом различных видов. Это трансмембранные белки-порины, формирующие в мембране гидрофильные поры диаметром примерно 1 нм, через которые проходят вода и гидрофильные низкомолекулярные вещества до 7кД (сахара, аминокислоты, небольшие олигосахариды и пептиды). Минорные белки НМ представлены гораздо большим числом видов. Их основные функции — транспортная (специфический транспорт в клетку железосодержащих соединений) и рецепторная;
8) Грам- бактерии менее чувствительны к пенициллину и лизоциму, чем Грам+.
Таким образом, у Грам- бактерий строение КС намного сложнее, чем у Грам+ (рис. 20 Б). В ее состав входит большее число разных химических макромолекул (табл. 3, 4).
Таблица 3
Различия между Грам+ и Грам- бактериями
| Признак | Грам+ | Грам- |
| Толщина КС | 20–60 нм | 10–20 нм |
| % содержание ПГ | 40–90% | 5–10% |
| Структура ПГ | Многослойный (10 слоев). Аминосахара (N–ацетилглюкозамин и N–ацетилмурамовая кислота) связаны гликозидными связями в гетерополимерную цепочку. Ацетилмурамовые кислоты гликановых цепей связаны между собой множественными связями через тетрапептиды разного строения. | Однослойный или двуслойный. Аминосахара (N–ацетилглюкозамин и N–ацетилмурамовая кислота) связаны гликозидными связями в гетерополимерную цепочку. Ацетилмурамовые кислоты гликановых цепей связаны между собой единичными связями через два однотипных тетрапептида. |
| Наличие ТК | + | - |
| Наличие периплазматического пространства между ЦПМ и ПГ | - | + |
| Наличие НМ | - | + |
| Выделение ферментов | В окружающую среду | В периплазматическое пространство |
| Окраска по Граму | Темно-фиолетовые | Розово-красные |
| Представители | Патогенные кокки, кроме гонокка и менингококка, бациллы, клостридии | Энтеробактерии, вибрионы, трепонемы |
| Спорообразование | Присутствует у бацилл и клостридий | Отсутствует |
Таблица 4
Химический состав КС Грам+ и Грам- бактерий
| Компоненты КС | Грам+ бактерии | Грам- бактерии | |
| внутренний слой (ПГ) | внешний слой (НМ) | ||
| Пептидогликан | + | + | – |
| Тейхоевые кислоты | + | – | – |
| Полисахариды | + | – | + |
| Белки | ± | – | + |
| Липиды | ± | – | + |
| Липополисахариды | – | – | + |
| Липопротеины | – | ± | + |
Прим.: (–) — отсутствуют, (+) — присутствуют, (±) — присутствуют не у всех видов
Функции клеточной стенки:
1. Обеспечиваетмеханическую защиту от воздействий окружающей среды.
Концентрация осмотически активных веществ (сахаров и солей) в клетке намного выше, чем в окружающей среде. Поэтому в клетке существует высокое осмотическое давление (у некоторых бактерий оно достигает 30 атмосфер). КС сдерживает это давление, предохраняет клетку от осмотического лизиса, дает возможность клетке существовать в гипотонических растворах. Если повысить осмотическое давление внешней среды (напр., путем добавления сахаров), вода будет оттягиваться из клетки.
2. Формообразующая. КС у бактерий не жесткая, как стальной панцирь, а эластичная, как кожаная покрышка футбольного мяча. Ригидность (упругость) и эластичность КС обеспечивает внутренний ПГ слой.
3. Транспортная: обеспечивает проникновение питательных веществ в клетку и удаление из нее продуктов метаболизма. КС проницаема для солей и других низкомолекулярных соединений благодаря наличию в НМ каналов (пор) для пассивного транспорта веществ и ионов, необходимых клетке. НМ также препятствует проникновению в клетку токсических веществ, поэтому Грам- бактерии (по сравнению с Грам+ бактериями) более устойчивы к действию некоторых ядов, химических веществ, ферментов и антибиотиков.
4. Содержит родо- и видоспецифические антигены (ЛПС Грам- бактерий, ТК Г+ бактерий).
Дата добавления: 2021-02-19; просмотров: 977;
Поиск по сайту
Узнать еще
- Выбор конструкции усиления и расчет усиленного простенка
- Генная и клеточная терапия моногенных и мультифакторных заболеваний
- Клеточная кооперация в иммунном ответе
- Клеточная стенка. Общая характеристика
- Клеточная структура животных тканей.
- Клеточная теория Шванна – Вирхова.
- Корпоративная сеть (КС) предприятия: назначение, структура и основные компоненты

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
