Транспорт веществ через плазматическую мембрану
Барьерно-транспортная функция поверхностного аппарата клетки обеспечивается избирательным переносом ионов, молекул и надмолекулярных структур в клетку и из нее. Транспорт через мембраны обеспечивает доставку питательных веществ и удаление конечных продуктов обмена из клетки, секрецию, создание ионных градиентов и трансмембранного потенциала, поддержание в клетке необходимых значений pH и др.
Механизмы транспорта веществ в клетку и из нее зависят от химической природы переносимого вещества и его концентрации по обе стороны клеточной мембраны, а также от размеров транспортируемых частиц. Малые молекулы и ионы транспортируются через мембрану путем пассивного или активного транспорта. Перенос макромолекул и крупных частиц осуществляется посредством транспорта в «мембранной упаковке», то есть за счет образования окруженных мембраной пузырьков.
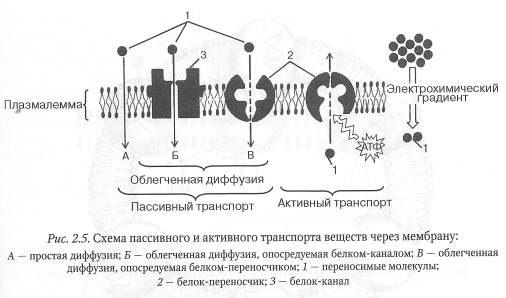
Пассивным транспортом называется перенос веществ через мембрану по градиенту их концентрации без затраты энергии. Такой транспорт осуществляется посредством двух основных механизмов: простой диффузии и облегченной диффузии.
Путем простой диффузии транспортируются малые полярные и неполярные молекулы, жирные кислоты и другие низкомолекулярные гидрофобные органические вещества. Транспорт молекул воды через мембрану, осуществляемый путем пассивной диффузии, получил название осмоса. Примером простой диффузии служит транспорт газов через плазматическую мембрану эндотелиальных клеток кровеносных капилляров в окружающую их тканевую жидкость и обратно.
Гидрофильные молекулы и ионы, не способные самостоятельно проходить через мембрану, транспортируются с помощью специфических мембранных транспортных белков. Такой механизм транспорта получил название облегченной диффузии.
Существуют два основных класса мембранных транспортных белков: белки-переносчики и белки-каналы. Молекулы переносимого вещества, связываясь с белком-переносчиком, вызывают его конформационные изменения, результатом чего служит перенос указанных молекул через мембрану. Облегченная диффузия отличается высокой избирательностью по отношению к транспортируемым веществам.
Белки-каналы формируют заполненные водой поры, пронизывающие липидный бислой. Когда эти поры открыты, неорганические ионы или молекулы транспортируемых веществ проходят сквозь них и таким образом переносятся через мембрану. Ионные каналы обеспечивают перенос примерно 106 ионов в секунду, что более чем в 100 раз превышает скорость транспорта, осуществляемого белками-переносчиками.
Большинство белков-каналов имеет «ворота», которые открываются на короткое время, а затем закрываются. В зависимости от природы канала «ворота» могут открываться в ответ на связывание сигнальных молекул (лиганд-зависимые воротные каналы), изменение мембранного потенциала (потенциал-зависимые воротные каналы) или механическую стимуляцию.
Активным транспортом называется перенос веществ через мембрану против их градиентов концентрации. Он осуществляется с помощью белков-переносчиков и требует затрат энергии, основным источником которой служит АТФ.
Примером активного транспорта, использующего энергию гидролиза АТФ для перекачки ионов Na+ и К+ через мембрану клетки, служит работа натриево-калиевого насоса, обеспечивающего создание мембранного потенциала на плазматической мембране клеток.
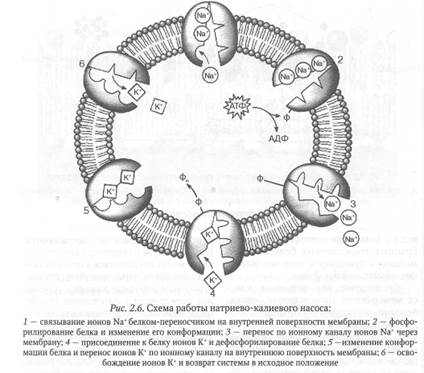
Насос образован встроенными в биологические мембраны специфическими белками-ферментами аденозинтрифосфатазами, катализирующими отщепление остатков фосфорной кислоты от молекулы АТФ. В состав АТФаз входят: ферментный центр, ионный канал и структурные элементы, препятствующие обратной утечке ионов в процессе работы насоса. На работу натриево-калиевого насоса расходуется более 1/3 АТФ, потребляемой клеткой.
В зависимости от способности транспортных белков переносить один или несколько видов молекул и ионов пассивный и активный транспорт подразделяются на унипорт и копорт, или сопряженный транспорт.
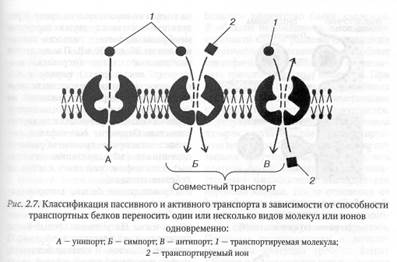
Унипорт — это транспорт, при котором белок-переносчик функционирует только в отношении молекул или ионов одного вида. При копорте, или сопряженном транспорте, белок-переносчик способен транспортировать одновременно два или более видов молекул или ионов. Такие белки-переносчики получили название копортеров, или сопряженных переносчиков. Различают два вида копорта: симпорт и антипорт. В случае симпорта молекулы или ионы транспортируются в одном направлении, а при антипорте — в противоположных направлениях. По принципу антипорта работает, например, натриевокалиевый насос, активно перекачивая ионы Na+ из клеток, а ионы К+ внутрь клеток против их электрохимических градиентов. Примером симпорта служит реабсорбция клетками почечных канальцев глюкозы и аминокислот из первичной мочи. В первичной моче концентрация Na+ всегда значительно выше, чем в цитоплазме клеток почечных канальцев, что обеспечивается работой натриево-калиевого насоса. Связывание глюкозы первичной мочи с сопряженным белком-переносчиком открывает Nа+-канал, что сопровождается переносом ионов Na+ из первичной мочи внутрь клетки по градиенту их концентрации, то есть путем пассивного транспорта. Поток ионов Na+, в свою очередь, вызывает изменения конформации белка-переносчика, результатом чего служит транспорт глюкозы в том же направлении, что и ионов Na+: из первичной мочи внутрь клетки. В данном случае для транспорта глюкозы, как можно убедиться, сопряженный переносчик использует энергию градиента ионов Na+, создаваемую работой натриево-калиевого насоса. Таким образом, работа натриево-калиевого насоса и сопряженного переносчика, использующего для транспорта глюкозы градиент ионов Na+, позволяет реабсорбировать практически всю глюкозу из первичной мочи и включить ее в общий метаболизм организма.
Благодаря избирательному транспорту заряженных ионов плазмалемма почти всех клеток несет на своей наружной стороне положительный, а на внутренней цитоплазматической стороне — отрицательный заряды. В результате этого между обеими сторонами мембраны создается разность потенциалов.
Формирование трансмембранного потенциала достигается в основном за счет работы встроенных в плазмалемму транспортных систем: натриевокалиевого насоса и белков-каналов для ионов К+.
Как отмечалось выше, в процессе работы натриево-калиевого насоса на каждые два поглощенных клеткой иона калия из нее выводится три иона натрия. В результате снаружи клеток создается избыток ионов Na+, а внутри — избыток ионов К+. Однако еще более значимый вклад в создание трансмембранного потенциала вносят калиевые каналы, которые в клетках, находящихся в состоянии покоя, всегда открыты. Благодаря этому ионы К+ выходят по градиенту концентрации из клетки во внеклеточную среду. В результате этого между двумя сторонами мембраны возникает разность потенциалов от 20 до 100 мВ. Плазмалемма возбудимых клеток (нервных, мышечных, секреторных) наряду с К+- каналами содержит многочисленные Nа+-каналы, которые открываются на короткое время при действии на клетку химических, электрических или других сигналов. Открытие Nа+-каналов вызывает изменение трансмембранного потенциала (деполяризацию мембраны) и специфический ответ клетки на действие сигнала.
Транспортные белки, которые генерируют разность потенциалов на мембране, называются электрогенными насосами. Натриево-калиевый насос служит главной электрогенной помпой клеток.
Транспорт в мембранной упаковкехарактеризуется тем, что транспортируемые вещества на определенных стадиях транспорта располагаются внутри мембранных пузырьков, то есть оказываются окруженными мембраной. В зависимости от того, в каком направлении переносятся вещества (в клетку или из нее), транспорт в мембранной упаковке подразделяется на эндоцитоз и экзоцитоз.
Эндоцитозом называется процесс поглощения клеткой макромолекул и более крупных частиц (вирусов, бактерий, фрагментов клеток). Эндоцитоз осуществляется путем фагоцитоза и пиноцитоза.
Фагоцитоз — процесс активного захвата и поглощения клеткой твердых микрочастиц, размер которых составляет более 1 мкм (бактерий, фрагментов клеток и др.). В ходе фагоцитоза клетка с помощью специальных рецепторов распознает специфические молекулярные группировки фагоцитируемой частицы.
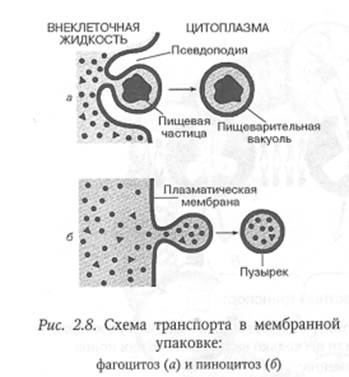
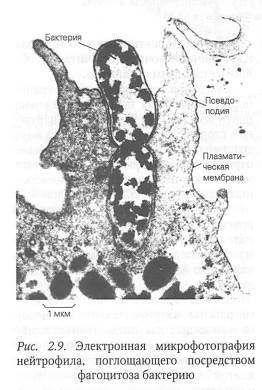
Затем в месте контакта частицы с мембраной клетки образуются выросты плазмалеммы — псевдоподии, которые обволакивают микрочастицу со всех сторон. В результате слияния псевдоподий такая частица оказывается заключенной внутри пузырька, окруженного мембраной, который называется фагосомой. Образование фагосом — энергозависимый процесс и протекает с участием актомиозиновой системы. Фагосома, погружаясь в цитоплазму, может сливаться с поздней эндосомой или лизосомой, в результате чего поглощенная клеткой органическая микрочастица, например бактериальная клетка, переваривается. У человека к фагоцитозу способны только немногие клетки: например, макрофаги соединительной ткани и лейкоциты крови. Эти клетки поглощают бактерии, а также разнообразные твердые частицы, попавшие в организм, и тем самым защищают его от болезнетворных микроорганизмов и посторонних частиц.
Пиноцитоз — поглощение клеткой жидкости в виде истинных и коллоидных растворов и суспензий. Этот процесс в общих чертах сходен с фагоцитозом: капля жидкости погружается в образовавшееся углубление клеточной мембраны, окружается ею и оказывается заключенной в пузырек диаметром 0,07—0,02 мкм, погруженный в гиалоплазму клетки.
Механизм пиноцитоза весьма сложен. Этот процесс осуществляется в специализированных областях поверхностного аппарата клетки, называемых окаймленными ямками, которые занимают около 2% клеточной поверхности. Окаймленные ямки представляют собой небольшие впячивания плазмалеммы, рядом с которыми в периферической гиалоплазме находится большое количество белка клатрина. В области окаймленных ямок на поверхности клеток располагаются также многочисленные рецепторы, способные специфически распознавать и связывать транспортируемые молекулы. При связывании рецепторами указанных молекул происходит полимеризация клатрина, и плазмалемма впячивается. В результате образуется окаймленный пузырек, несущий в себе транспортируемые молекулы. Свое название такие пузырьки получили благодаря тому, что клатрин на их поверхности под электронным микроскопом выглядит как неровная каемка. После отделения от плазмалеммы окаймленные пузырьки теряют клатрин и приобретают способность сливаться с другими пузырьками. Процессы полимеризации и деполимеризации клатрина требуют затрат энергии и блокируются при недостатке АТФ.
Пиноцитоз, благодаря высокой концентрации рецепторов в окаймленных ямках, обеспечивает избирательность и эффективность транспорта специфических молекул. Например, концентрация молекул транспортируемых веществ в окаймленных ямках в 1000 раз превышает концентрацию их в окружающей среде. Пиноцитоз — основной способ транспорта в клетку белков, липидов и гликопротеинов. Посредством пиноцитоза клетка поглощает за сутки количество жидкости, равное своему объему.
Экзоцитоз — процесс выведения веществ из клетки. Вещества, подлежащие выведению из клетки, сначала заключаются в транспортные пузырьки, наружная поверхность которых, как правило, покрыта белком клатрином, затем такие пузырьки направляются к клеточной мембране. Здесь мембрана пузырьков сливается с плазмалеммой, а содержимое их изливается за пределы клетки либо, сохраняя связь с плазмалеммой, включается в гликокаликс.
Существуют два типа экзоцитоза: конститутивный (основной) и регулируемый.
Конститутивный экзоцитоз непрерывно протекает во всех клетках организма. Он служит основным механизмом выведения из клетки продуктов метаболизма и постоянного восстановления клеточной мембраны.
Регулируемый экзоцитоз осуществляется лишь в специальных клетках, выполняющих секреторную функцию. Выделяемый секрет накапливается в секреторных пузырьках, а экзоцитоз происходит только после получения клеткой соответствующего химического или электрического сигнала. Например, β-клетки островков Лангерганса пожелудочной железы выделяют свой секрет в кровь лишь при повышении в крови концентрации глюкозы.
В ходе экзоцитоза сформировавшиеся в цитоплазме секреторные пузырьки обычно направляются к специализированным участкам поверхностного аппарата, содержащим большое количество фузионных белков или белков слияния. При взаимодействии белков слияния плазмалеммы и секреторного пузырька образуется фузионная пора, соединяющая полость пузырька с внеклеточной средой. При этом активируется актомиозиновая система, в результате чего содержимое пузырька изливается из него за пределы клетки. Таким образом, при индуцируемом экзоцитозе энергия требуется не только для транспорта секреторных пузырьков к плазмалемме, но и для процесса секреции.
Трансцитоз, или рекреция, — это транспорт, при котором происходит перенос отдельных молекул через клетку. Указанный вид транспорта достигается за счет сочетания эндо- и экзоцитоза. Примером трансцитоза служит транспорт веществ через клетки сосудистых стенок капилляров человека, который может осуществляться как в одном, так и в другом направлениях.
Дата добавления: 2016-10-18; просмотров: 19553;
Поиск по сайту
Узнать еще
- I.2.2 ТЕПЛОЁМКОСТЬ ВЕЩЕСТВА
- II. Поведение веществ в магнитном поле.
- III. Светофоры на железнодорожном транспорте
- IV. Сигналы ограждения на железнодорожном транспорте
- IX. Звуковые сигналы на железнодорожном транспорте
- UMTS интерфейсы транспортной сети
- V. Ручные сигналы на железнодорожном транспорте
- VI. Пектиновые вещества и их роль в пищевой промышленности.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
