Класс земноводные или Амфибии – Amphibia
Земноводные – самый малочисленный класс современных позвоночных: в трех отрядах: бесхвостые – Anura (Ecaudata), хвостатые – Urodella (Caudata), безногие – Apoda, около 2500 видов.
Амфибии – первые наземные позвоночные, еще сохранившие значительные связи с водной средой. У большинства видов яйца (икра) лишены плотных оболочек и могут развиваться только в воде. Вылупившиеся из яиц личинки ведут водный образ жизни и лишь затем совершают метаморфоз (превращение), во время которого формируются особенности взрослых, ведущих наземный образ жизни.
Для взрослых земноводных характерны парные конечности с шарнирными суставами. Череп двумя затылочными мыщелками подвижно сочленяется с шейным позвонком, небно-квадратный хрящ срастается с мозговой коробкой (аутостилический череп), а верхний элемент подъязычной дуги (подвесок) превращается в косточку среднего уха – стремечко. Тазовый пояс причленяется к поперечным отросткам крестцового позвонка. Формируется два круга кровообращения, полностью не разобщенные; в сердце два предсердия, но один желудочек. Глаза имеют подвижные веки. Органы боковой линии у взрослых обычно исчезают. Передний мозг увеличивается и разделяется на два полушария; в его крыше есть скопления нервных клеток. Средний уровень метаболизма амфибий заметно выше уровня рыб. Наряду с этим земноводные сохранили и признаки водных позвоночных. Проницаемая для воды и газов голая кожа имеет большое количество слизистых желез. Органами выделения служат туловищные (мезонефрические) почки и кожа. Температура тела зависит от температуры окружающей среды и лишь слегка превышает последнюю (пойкилотермия).
Центральная нервная система и органы чувств.
Переход к наземному образу жизни сопровождался преобразованием центральной нервной системы и органов чувств. Относительные размеры головного мозга земноводных по сравнению с рыбами заметно не возрастают. У бесхвостых несколько крупнее, чем у хвостатых. Вес головного мозга от массы тела составляет у современных хрящевых рыб 0,06-0,44%, у костных рыб 0,02-0,94%, у хвостатых земноводных 0,29-0,36%, у бесхвостых 0,50-0,73%. Следует отметить, что у современных земноводных мозг, вероятно, несколько уменьшен по сравнению с мозгом предков – стегоцефалов (об этом свидетельствует сопоставление размеров мозговых черепов).
У современных земноводных заметно увеличиваются относительные размеры переднего мозга, разделившегося на два полушария с самостоятельной полостью (боковым желудочком) в каждом из них. Скопления нервныхклеток образуют не только полосатые тела в дне боковых желудочков, но и тонкий слой в крыше полушарий (первичный мозговой свод) – архипаллиум (из современных рыб он есть у двоякодышащих). Обонятельные доли слабо отграничены от полушарий. Промежуточный мозг лишьслегка прикрыт соседними отделами. Сверху на нем расположен эпифиз. От дна промежуточного мозга отходит воронка, к которой прилегает хорошо развитый гипофиз. Мозжечок мал и имеет вид небольшого валика, лежащего за средним мозгом по переднему краю ромбовидной ямки – полости четвертого желудочка. От головного мозга земноводных, как у рыб, отходят 10 пар головных нервов; XII пара (подъязычный нерв) отходит за пределами черепной коробки, а добавочный нерв (XI пара) не развивается.
Развитие архипаллиума, сопровождающееся усилением связей с промежуточным и особенно средним мозгом, приводит к тому, что ассоциативная деятельность, регулирующая поведение, осуществляется не только продолговатым и средним мозгом, но и полушариями переднего мозга. У хвостатых земноводных уровень нервной деятельности ниже, нежели у бесхвостых; это связано с относительно меньшей величиной головного мозга и тонкостью архипаллиума (около 0,2 мм против 0,6-0,8 мм у бесхвостых). Слабое развитие мозжечка у всех земноводных соответствует простоте (стереотипности) движений (рис. 50).
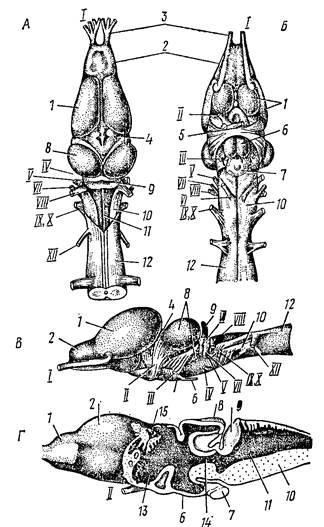

Рис. 50. Головной мозг лягушки : А – сверху, Б – снизу, В – сбоку, Г – в продолном разрезе (по Н. П. Наумову, Н. Н. Карташеву, 1979).
Слегка сплющенный спинной мозг имеет плечевое и поясничное утолщение, связанные с отхождением мощных нервных сплетений, иннервирующих передние и задние конечности. По сравнению с рыбами усиливается разделение серого и белого вещества, т.е. усложняются проводящие нервные тракты. Спинномозговых нервов у бесхвостых 10 пар, у хвостатых – в зависимости от числа позвонков – несколько десятков пар. Симпатическая нервная система у земноводных представлена двумя стволами, лежащими по бокам брюшной стороны позвоночною столба. Ганглии этих стволов соединены со спинномозговыми нервами.
Органы чувств обеспечивают ориентировку земноводных в воде и на суше. У личинок и у ведущих водный образ жизни взрослых земноводных важную роль играют органы боковой линии (сейсмосенсорная система) осязание, терморецепция, вкус, слух и зрение. У видов с преимущественно наземным образом жизни основную роль в ориентации играет зрение. Органы боковой линии есть у всех личинок и у взрослых с водным образом жизни. Они разбросаны по всему телу (более густо на голове) и в отличие от рыб лежат на поверхности кожи. В поверхностных слоях кожи разбросаны осязательные тельца (скопления чувствующих клеток с подходящими к ним нервами). У всех земноводных в эпидермальном слое кожи имеются свободные окончания чувствующих нервов. Они воспринимают температурные, болевые и тактильные ощущения. Часть из них, видимо реагирует на изменения влажности и, возможно, на изменение химизма окружающей среды. В ротовой полости и на языке имеются скопления чувствующих клеток, оплетенные нервными окончаниями. Однако они, видимо, выполняют не функцию ''вкусовых" рецепторов, а служат органами осязания, позволяющими ощущать положение пищевого объекта в ротовой полости. О слабом развитии вкуса у земноводных свидетельствует поедание ими насекомых с резким запахом и едкими выделениями (муравьи, клопы, жужелицы и др.).
Обоняние, видимо, играет заметную роль в жизни амфибий. Обонятельные мешки парные. Наружные ноздри открываются и закрываются действием специальных мышц. Внутренними ноздрями (хоанами) каждый мешок сообщается с ротовой полостью. Поверхность обонятельных мешков увеличивается продольной складчатостью их стенок и боковыми выпячиваниями. Трубчатые железы стенок выделяют секрет, смачивающий слизистую обонятельных мешков. Лишь часть стенок обонятельных мешков выстлана специальным обонятельным эпителием, к клеткам которого подходят окончания обонятельного нерва. Объем обонятельных мешков и их площадь, занятая обонятельным эпителием, особенно велики у безногих (червяг) и части бесхвостых (жабы, некоторые квакши). Орган обоняния функционирует только в воздушной среде, в воде наружные ноздри закрыты. Роль обоняния в ориентации и поисках пищи велика у роющих червяг. Хвостатые и бесхвостые земноводные распознают запах мест обитания, запах "своего" или "чужого" вида, запах пищи. Чувствительность обоняния меняется в разные сезоны; особенно высока она весной. На запахи у амфибий удается выработать условные рефлексы.
У всех земноводных в области хоан (внутренние ноздри) образуются небольшие слепые углубления, стенки которых выстланы чувствующим эпителием, иннервируемым ветвями обонятельного нерва. Полость этих углублений заполнена секретом специальных желез. Эти органы называют якобсоновыми и полагают, что они служат для восприятия запаха пищи, находящейся в ротовой полости. У червяг в ямке на голове находится подвижное щупальце, которое постоянно высовывают, как бы ощупывая пространство около головы, полагают, что оно выполняет функцию не только осязания, но и обоняния. Органы зрения хорошо развиты у подавляющего большинства земноводных. Лишь у живущих в почве червяг и постоянных обитателей подземных водоемов – европейского протея, подземной саламандры и нескольких других видов – маленькие глаза слегка просвечивают сквозь кожу или не видны. По сравнению с рыбами роговица глаз амфибий более выпукла, а хрусталик имеет форму двояковыпуклой линзы с более плоской передней поверхностью. Аккомодация осуществляется лишь перемещением хрусталика при помощи мышечных волокон ресничного тела. Глаза личинок, как и рыб, не имеют век. Во время метаморфоза образуются подвижные веки – верхнее и нижнее, и мигательная перепонка (обособляется от нижнего века). Секрет желез внутренней поверхности век и мигательной перепонки предохраняет роговицу от высыхания; при движении век с поверхности глаза удаляются осевшие посторонние частицы.
В сетчатке есть палочки и колбочки; у видов с сумеречной и ночной активностью первые преобладают. Общее число фоторецепторных клеток у хвостатых амфибий колеблется в пределах 30-80 тыс. на 1 мм2 сетчатки, а у бесхвостых – до 400-680 тыс. У многих земноводных развито цветовосприятие.
Показано, что цветоразличие обеспечивается в так называемом ядре Беллоци (промежуточный мозг), тогда как основная информация поступает в зрительную кору. В сетчатке группы рецепторов (палочки и колбочки) связаны с биполярными клетками через поперечные и амакриновые нейроны; группы биполяров передают полученную информацию на детекоры – ганглиозные клетки. Выяснено, что ганглиозные клетки сетчатки лягушек представлены несколькими функциональными типами. Одни реагируют на попавшие в поле зрения мелкие округлые предметы – пищу (детекторы формы); вторые – контрастируют изображение, выделяя его на общем фоне (детекторы контраста); третьи (детекторы движения) – реагируют на перемещение "пищи", а четвертые – на быстрое и общее затемнение поля зрения (расценивается как сигнал опасности – приближение врага). Существуют и "дирекционные" нейроны, регистрирующие направление движешь "пищи"; они связаны с базальным ядром промежуточного мозга. Таким, образом, первичная обработка (классификация) зрительных сигналов в отличие от других позвоночных у амфибий происходит уже в сетчатке. Собираемая информация небогата. Неподвижные земноводные воспринимают лишь движения мелких объектов или приближение врага; все остальное представляется им индифферентным ''серым фоном". При движении они начинают различать и неподвижные объекты. Благодаря положению глаз у многих земноводных общее поле зрения равно 360° при значительном секторе бинокулярного зрения, позволяющего оценивать расстояние до перемещающегося пищевого объекта, что дает возможность успешно ловить мелкую подвижную добычу. На основе изучения механизмов зрения лягушки созданы фототехнические устройства, распознающие мелкие объекты.
Орган слуха в связи с земноводным образом жизни существенно усложнился. Заключенный в капсулу внутреннего уха перепончатый лабиринт у земноводных по сравнению с рыбами, изменился мало: увеличились размеры полого выступа (lagena) и возросла площадь чувствующих полей. Возник новый отдел – среднее ухо, в котором расположен аппарат, усиливающий воспринятые звуковые колебания. Необходимость его связана со слабой звукопроводимостью воздуха по сравнению с водой. Полость среднего уха образовалась из рудимента жаберной щели. Она располагается между челюстной и подъязычной дугами и сохранившийся у многих рыб в виде брызгальца – канала, соединяющего полость глотки с внешней средой. Наружное отверстие полости среднего уха затянуто тонкой упругой барабанной перепонкой. В полости находится палочковидная косточка – стремечко (stapes), одним концом упирающаяся в барабанную перепонку, а другим – в овальное окно, которое закрыто меньшей по сравнению с барабанной перепончатой перегородкой. Это и позволяет усиливать слабые звуковые колебания. Сравнительно-анатомические и палеонтологические данные показывают, что стремечко образовалось из гиомандибуляре или подвеска – верхнего элемента подъязычной дуги. Узкий канал – евстахиева труба, соединяя полость среднего уха с ротовой полостью, выравнивает давление и предотвращает разрывы барабанной перепонки при сильных звуках.
Наряду с системой – среднее и внутреннее ухо, обеспечивающей слух в воздушной среде, у земноводных сохраняются механизмы слуха в воде. Звукопроводимость тела амфибий, подобно рыбам, близка к водной среде, что позволяет принимать звуки непосредственно на мембрану овального окна. Этому способствуют поверхностные вены и эндолимфатический проток. Звуки в воде принимаются так называемым macula sacculi – гомологом звукорецептора рыб, а звуки в воздухе, переданные стремечком с барабанной перепонки воспринимаются звуковым рецептором наземных позвоночных. Существование двух слуховых систем у амфибий – приспособление к жизни в двух средах – водной и наземной.
У немногих бесхвостых (жерлянки, чесночницы), у всех хвостатых и безногих земноводных полость среднего уха и барабанная перепонка вторично редуцируются. В воде эти животные используют описанный выше механизм звуковосприятий, а возможности их слуха в воздушной среде, видимо, невелики. Червяги способны улавливать распространяющиеся по земле звуки через костный путь: квадратная кость – стремечко. В ослабленной форме такая звуковая передача возможна и для хвостатых земноводных. Таким образом, земноводные обладают достаточно широкими возможностями слуха в воде, а бесхвостые – и в воздухе. Последние воспринимают звуки от 30 до 15000 Гц. Наличие в гортанно-трахейной полости голосовых складок позволяет бесхвостым земноводным издавать видоспецифические звуки: их громкость усиливается имеющимися у части видов резонаторами – специальными полостями в углах рта, которые при кваканье раздуваются. Звуковые сигналы амфибий преимущественно обслуживают размножение; они помогают встрече самцов и самок своего вида и различию чужих видов. Некоторые звуки служат сигналом опасности.
Дата добавления: 2022-04-12; просмотров: 247;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
