Класс Костные рыбы – Osteichthyes
Населяют практически все водоемы земного шара: толщу морей и океанов, разнообразные пресные водоемы, даже пересыхающие и пещерные. По обилию видов (около 20 тыс.) – самый многочисленный класс позвоночных животных.
Класс разделяется на Подкласс Лопастеперые рыбы – Sarcopterygii. Надотряд Кистеперые рыбы – Grossopterygimorpha, отряд Целакантообразные – Coelacanthiformes (латимерия). Надотряд Двоякодышащие рыбы – Dipneustomorpha. Отряд Однолегочные – Ceratodiformes (цератод). Отряд Двулегочнообразные – Lepidosireniformes (лепидосирен).
Подкласс Лучеперые рыбы – Actinopterygii включает надотряд Ганоидные (Ganoidomorpha) с отрядами: осетрообразные, многоперообразные, амиеобразные, панцирникообразные. Последние 2 отряда часто объединяют в группу костные ганоиды – Holostei. Все остальные отряды, примерно 30-40 отрядов, объединяют в группу костистые рыбы – Teleostei.
У костистых рыб относительные размеры головного мозга в большинстве случаев крупнее, чем у хрящевых рыб. В то же время функции его отделов в обоих классах одинаковы. Передний мозг костистых рыб относительно невелик. Главную его массу составляют полосатые тела. Нервные клетки в крыше переднего мозга отсутствуют или образуют лишь незначительные скопления. Промежуточный мозг довольно велик, на его спинной стороне образуется хорошо развитый эпифиз, а на брюшной стороне – гипофиз. Зрительные нервы образуют отчетливый перекрест (хиазму). Средний мозг у большинства костных рыб по размерам заметно превышает остальные отделы мозга и прикрывает промежуточный мозг. Сверху он подразделен на две зрительные доли, в которых оканчиваются волокна зрительного нерва; там проходит обработка зрительных восприятий. В брюшной части среднего мозга сосредоточены центры связи с мозжечком, продолговатым и спинным мозгом, в которых осуществляется суммация и сопоставление восприятий. У подвижных видов рыб мозжечок хорошо развит, он частично налегает на средний мозг и почти прикрывает продолговатый мозг. У малоподвижных рыб размеры мозжечка уменьшаются. Осуществляя взаимодействие восходящих путей спинного и продолговатого мозга и нисходящих путей среднего мозга. Мозжечок обеспечивает контроль над тонусом соматической мускулатуры, координацией движения и сохранением равновесия. Продолговатый мозг по форме, относительным размерам и функции сходен с продолговатым мозгом хрящевыхрыб (рис. 49).
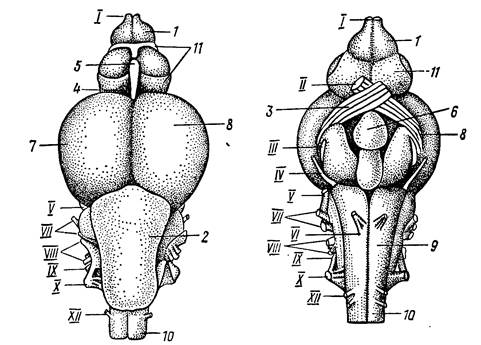
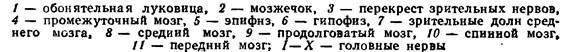
Рис. 49. Головной мозг форели сверху и снизу (по Н. П. Наумову, Н. Н. Карташеву, 1979).
У двоякодышащих, кистеперых и многоперообразных рыб пропорции отделов головного мозга отличаются от описанных выше и характерных для остальных костных рыб. Передний мозг у них крупнее остальных отделов; он разделяется на правое и левое полушария. В области желудочков нервные клетки подстилают крышу полушарий. Средний мозг и мозжечок имеют относительно небольшие размеры. Эти отличия, видимо, связаны с большей ролью обоняния, по сравнению со зрением, и малой подвижностью этих рыб.
Как и у хрящевых, от головного мозга костных рыб отходят 10 пар головных нервов.
Спинной мозг костных рыб по строению сходен со спинным мозгом хрящевых рыб. Он лежит в канале, образованном верхними дугами позвонков. Автономность функционирования спинного мозга у костных рыб выражена слабее, чем у хрящевых рыб.
Как и у хрящевых, в ориентации и общении костных рыб особенно, важную роль играют органы химического чувства (обоняние и вкус). Оба канала получения химической информации взаимодействуют, дополняя друг друга.
Обонятельные мешки костных рыб имеют внутри хорошо развитые складки обонятельного эпителия. Кожистый клапан разделяет ноздри надваотверстия: у плывущей рыбы вода заходит в обонятельный мешок через переднее и выходит через заднее носовое отверстие. У части рыб (плотва, угри и др.) ток воды через обонятельный мешок усиливается мерцанием ресничек, усеивающих его внутреннюю поверхность. Хотя обонятельные доли и весь передний мозг у большинства костных рыб относительно невелик (сильнее развиты у кистеперых, двоякодышащих и многоперообразных), пороги обонятельной чувствительности очень низки. Морские налимы, сидевшие в бассейне емкостью 300 л, тотчас реагировали на прибавление 2,5 л воды из 12-литровой банки, в которой лишь 5 минут находилась одна рыбка атеринка. Еще ниже порог чувствительности у угря: запах воспринимается при наличии 15-50 тыс. молекул в 1 л (примерно соответствует чутью собаки). Биологическое значение обоняния костных рыб многообразно. Эксперименты показали, что рыбы хорошо распознают запахи не только чужих видов, но и различных особей своего вида. Это позволяет стайным рыбам не терять свою стаю, одиночным – находить партнера при размножении. Распознавание запаха чужого вида позволяет уклоняться от нападения хищника или уловить приближение добычи. У многих рыб, особенно стайных, в особых бокаловидных клетках кожи содержится так называемое "вещество страха" или "опасности". При ранении кожи она попадает в воду и воспринимается другими рыбами как сигнал опасности. Под действием такого химического сигнала стая уплотняется, что затрудняет нападение хищника, или, наоборот, рассеивается, ирыбы затаиваются. Велико значение обоняния и "химической" памяти у мигрирующих рыб. Так, тихоокеанские лососи выклевываются из икры в реках, проводят там несколько месяцев на стадии личинки и малька, а затем скатываются в море и кочуют. Через несколько лет, достигнув половой зрелости, рыбы мигрируют на нерест, проплывают сотни километров, и находят родные реки по запаху их вод. Меченьем личинок было установлено, что из 13 тысяч, выловленных на нерестилищах рыб, 34%вошли точно в те же реки и ручьи, где они выклюнулись из икры, 65% – в соседние, и лишь 1% рыб был пойман на значительном удалении от мест их мечения.
При поисках и распознавании пищи важное значение имеет вкус. Вкусовые почки (скопление чувствующих клеток с оканчивающимися на них веточками головных нервов) расположены в слизистой оболочке poтовой области (щука, колюшки). У многих рыб рассеяны по всей поверхности тела, концентрируясь на голове, усиках, иногда на удлиненных лучах плавников. Экспериментально показано, что рыбы различают сладкое и соленое, кислое, горькое. Нитевидные грудные плавники некоторых придонных рыб высокочувствительны к механическим раздражениям и экстрактам из различных морских беспозвоночных. Острота вкуса связана с экологическими особенностями вида: пищевой специализацией, типом местообитания, степенью развития других рецепторов. Слепая пещерная рыбка распознает раствор глюкозы при концентрации 0,005%.
У костных рыб, как и у хрящевых хорошо развиты органы боковой линии (сейсмосенсорные органы). У большинства костных рыб канал боковой линии проходит в толще кожи вдоль средней линии тела, а на голове сильно ветвится с образованием сложной сети. Многочисленные мелкие отверстия, пронизывающие чешуи, сообщают полость канала с внешней средой. Расположенные на стенках канала скопления снабженных ресничками чувствующих клеток иннервируются боковой ветвью блуждающего нерва. Органы боковой линии воспринимают даже слабые перемещения частиц воды и инфразвуковые колебания. С помощью этих органов рыбы ориентируются в потоках воды, воспринимают приближение или удаление добычи, хищника или партнера по стае, избегают столкновения с подводными предметами. Тонкость такой ориентации настолько велика, что ослепленные рыбы плавают с обычной скоростью, не натыкаясь на препятствия; они поддерживают строй в стае и успешно схватывают плавающую добычу.
К механорецепторам относятся и органы осязания. Они представлены расположенными в коже осязательными тельцами (скоплениями чувствующих клеток), рассеянными по всей поверхности тела и обычно более многочисленными на губах, усиках и лучах плавников. С их помощью рыбы воспринимают раздражения от прикосновения твердых тел. Teмпература воды воспринимается с помощью терморецепторов, рассеянных в поверхностных слоях кожи свободных нервных окончаний, видимо, различающихся по реакциям на разную температуру. Костные рыбы способны различать перепады температуры в 0,4°С.
Рыбы способны улавливать изменения магнитного и электрического поля. При низком напряжении электрического поля обычно ориентируют тело по силовым линиям, при увеличении напряжения двигаются к аноду, при возрастании напряжения наступает гальванонаркоз. Сильные электрические поля, возникающие у линий высокого напряжения, оказывают на поведение рыб угнетающее влияние. Электрические поля образуются вокруг тела любой рыбы, но они настолько слабы, что рецепторами не воспринимаются. Когда неэлектрические рыбы держатся группами (стаями), происходит суммация их индивидуальных электрических полей и образуется электрическое поле стаи, достаточное для восприятия отдельными членами. Оно играет важную роль в согласовании поведения отдельных членов стаи и их пространственной ориентации. Рецепторы электромагнитного чувства расположены преимущественно на голове.
Среди костистых рыб есть виды, имеющие специальные электрические органы, у отдельных видов – большой мощности. В большинстве случаев они представлены участками преобразованной поперечнополосатой мускулатуры. У рыб из отряда клюворылообразных, населяющих мутные водоемы тропической Африки, небольшие электрические органы расположены на хвостовом стебле. Их разряды создают вокруг рыбы электрическое поле, изменение которого позволяет рыбам оценивать характер предметов на расстоянии в несколько метров и тем самым обеспечивать ориентировку и поиск добычи в мутной воде. Электрические поля слабо электрических рыб служат средством разграничения кормовых участков, предотвращая вторжение на занятую территорию. В спокойном состоянии мормир генерирует 1 импульс в 1 секунду, а при беспокойстве – до 30 импульсов в 1 секунду. Плавающий гимнарх генерирует до 300 импульсов в 1 секунду напряжением в несколько вольт. Рецепторами служат кожные нервножелезистые клетки, расположенные преимущественно на спине и вканалах боковой линии.
У части населяющих прибрежные тропические воды рыб – звездочетов на голове есть небольшие электрические органы. У одного вида звездочетов электрический орган, образованный преобразованными мышцами глаз, способен генерировать разряды напряжением 50 вольт. Звездочет полузакопавшись в грунт лежит на дне, когда мелкая рыбешка подплывает близко, следует разряд, и оглушенная добыча падает прямо в открывшийся рот хищника. У электрических угрей, населяющих пресные водоемы Южной Америки, расположенные по бокам тела электрические органы генерируют до 20-50 импульсов в 1 секунду и создают электрическое поле, которое обеспечивает ориентировку рыбы (глаза у них очень малы).У электрического угря (Electrophorus electricus), достигающего 2 м длины, большие лентовидные электрические органы помимо низковольтных "ориентированных" импульсов генерируют сильные разряды. У живущего в водоемах Западной Африки электрического сома электрическийорган в виде сплошного студенистого подкожного слоя окружает все туловище, он используется как орган ориентации, защиты и нападения. Зрение рыб – важное средство ближней ориентации и связи рыб в стаях.Глаза костных рыб имеют уплощенную роговицу и шаровидный хрусталик. Подвижных век нет. Относительные размеры глаз в пределах класса варьируют. У рыб, живущих в условиях плохой освещенности (мутная вода, большие глубины и т.д.), глаза либо относительно очень крупны (у некоторых приобретают «телескопическую форму»), либо оченьмалы, рудиментарны, а у некоторых пещерных рыб скрыты под коржей. В последнем случае сильно развиты органы рецепции (боковая линия,обоняние, электрические органы и т.д.). В зависимости от образа жизни и пищевой специализации положение глаз изменчиво: у донных видов они смещены к верхней стороне головы, у камбалы оба глаза лежат на одной стороне головы. У выползающего на берег илистого прыгуна крупные глаза как бы приподняты над головой и очень подвижны. У охотящейся на поверхности воды четырехглазки крупные глаза с двойным зрачком и овальным хрусталиком позволяют одновременно видеть и в воздухе, и в воде. У большинства видов глаза размещены по бокам головы. Зрение преимущественно монокулярное: при поле зрения каждого глаза в 160-170°по горизонтали и около 150° по вертикали, поле бинокулярного зрения составляет всего 20-30°.
В хрящевой склере иногда образуются мелкие окостенения. Наружная часть сосудистой оболочки обособляется в серебристую оболочку. У многих хрящевых и костных рыб на внутренней поверхности сосудистой оболочки образуется зеркальце или тапетум – слой плоских клеток, наполненных блестящими кристалликами гуанина. Он отражает прошедшие через сетчатку световые лучи, что усиливает возможности зрения при слабой освещенности. Тонкий вырост сосудистой оболочки – серповидный отросток пронизывает сетчатку и стекловидное тело, прикрепляясь к хрусталику. При сокращении серповидного отростка хрусталик несколько сдвигается вглубь глаза, и так осуществляется его аккомодация (настройка на резкость). Тонкий пигментный слой у видов с хорошо развитым тапетумом выражен слабо.
У большинства костных рыб сетчатка смешанная: содержит и палочки, и колбочки. При сильном свете увеличивающиеся отростки пигментных клеток закрывают палочки, а колбочки подтягиваются к своим ядрам. При перемещении рыбы в темноту сокращаются пигментные клетки и прикрывают приблизившиеся к ним концевые членики колбочек, палочки подтягиваются к ядрам. Подобная реакция (свойственная и всем остальным позвоночным) обеспечивает адаптацию зрения к меняющейся освещенности. Количество палочек и колбочек и их соотношение в сетчатке разных видов рыб сильно варьирует.
Рыбы воспринимают световые волны длиной в 400-750 нм. Максимумы световой чувствительности у пресноводных рыб по сравнению с морскими несколько сдвинуты в длинноволновую часть спектра. Большинство костных рыб имеют цветное зрение (некоторые, видимо, примерно в таких же пределах, как и человек). Глаза используются как рецепторы ближней ориентации (не далее 10-15 м). При равных условиях крупные рыбы имеют большую остроту зрения. Зрение играет важную роль при поисках пищи, уходе от опасности и при внутривидовом общении: при контактах в стае, поисках партнера для размножения, у некоторых рыб – при согласованном поведении молоди и родителей. Цветное зрение позволяет использовать особенности окраски для распознавания особей своего вида и в некоторых случаях их физиологического состояния.
Орган слуха и равновесия костных рыб, как и хрящевых, представлен только внутренним ухом, заключенным в хрящевую капсулу, наружные стенки которой окостеневают. Перепончатый лабиринт, или собственно внутреннее ухо, образовано лежащими во взаимоперпендикулярных плоскостях тремя хорошо развитыми полукружными каналами, отходящими от овального мешочка (вестибулярный аппарат или орган равновесия), лежащий ниже круглый мешочек имеет более или менее четко выраженный полый вырост (лагену) и служит собственно органом слуха. Отходящий от круглого мешочка тонкий эндолимфатический проток заканчивается слепо. В полости лагены, круглого и овального мешочков лежат отолиты, или слуховые камешки. Они образованы кристалликами углекислой извести, скрепленными органическим веществом (по их слоистости, видной на шлифах, у многих видов можно определить возраст). У части костных рыб возникает связь плавательного пузыря с перепончатым лабиринтом: слепые отростки плавательного пузыря примыкают к затянутым перепонкой окошкам перилимфатической полости или образуется Веберов аппарат – система косточек, соединяющих стенку плавательного пузыря с перилимфатической полостью внутреннего уха. Благодаря этому перепончатый лабиринт служит рецептором, улавливающим изменение давления в плавательном пузыре, а плавательный пузырь выполняет функцию резонатора и тем самым увеличивает остроту слуха.
Костные рыбы воспринимают звуковые волны частотой от 16 до 12 000 Гц. Рецептором служат чувствительные поля лагены и круглого мешочка, а возможно, и овального мешочка. Низкочастотные звуки не выше 500-600 Гц, видимо, могут восприниматься и органами боковой линии. Следует напомнить, что в воде звуки распространяются со значительно большей скоростью, чем в воздухе (около 1500 м/с против 330 м/с), и на большие расстояния. Поэтому звуковая ориентация для животных, в том числе и для рыб, очень важна. Звукопроводимость тканей тела рыб близка звукопроводимости воды и поэтому восприятие звуков возможно при относительно простом строении органа слуха.
В последние десятилетия выяснилось, что ходячее выражение "нем как рыба" не соответствует действительности. Рыбы издают разнообразие звуки, воспринимаемые особями того же и других видов, как сигналы определенного значения. Механические или специфические звуки, возникающие при различных действиях рыбы (плавании, дыхании, еде), тоже имеют сигнальное значение. Звуки челюстей при схватывании добычи и перетирании пищи привлекают других особей своего вида или хищников, звуки питания хищных рыб вызывают оборонительную реакцию у мирных рыб: бегство, затаивание, уплотнение стаи и т.п. Привлекает хищников и звук трепетания или "крики боли" схваченной рыбы. Более определенное видоспецифическое значение имеют звуки, специально производимые рыбой в определенной ситуации: при встрече партнеров в период, размножения (нерестовые сигналы, включающие призыв особей другого пола, опознавание и стимуляцию выделения половых продуктов), сигналу предупреждения и угрозы при защите отложенной икры или охране своей территории и т.п. Эти специфические сигналы по звучанию могут быть очень разнообразными. Скрипучие и скрежечущие звуки издаются при произвольном трении друг о друга свободных костных лучей плавников костей челюстей и жаберной крышки, пояса грудных плавников и т.д.
Особенно разнообразны звуки, издаваемые с помощью плавательного пузыря: они напоминают барабанный бой, хлопки, свист, ворчание, кудахтанье, жужжание, стоны и т.п. Звуковые вибрации стенок плавательного пузыря могут вызывать колебания косточек веберова аппарата, костей пояса грудных плавников или ритмичные удары плавниковых лучей по телу. Частоты издаваемых рыбами звуков лежат в пределах 20-12000 Гц,т.е. в пределах чувствительности их звукового аппарата. Наиболее разнообразны акустические сигналы свойственные сумеречным и придонным рыбам, рыбам со сложной популяционной структурой.
Таким образом, костные рыбы имеют разнообразные органы чувств. При ориентировке и при поисках пищи почти всегда используются многие рецепторы. Так, нарушения положения тела, например, при ударе волны, воспринимаются глазами, боковой линией, полукружными каналами к чувствительными полями круглого и овального мешочков (участками, к которым прилегают отолиты), плавательным пузырем и осязательными тельцами. При отыскивании подвижной добычи используется зрение, органы боковой линии и электрические органы. Обоняние и осязание помогают находить неподвижную пищу, а захват и проглатывание пищи проходит под контролем зрения и вкусовых и осязательных телец, расположенных на усиках и в ротовой полости.
Дата добавления: 2022-04-12; просмотров: 599;
Поиск по сайту

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
