Обмен веществ у бактерии
При сравнительно бедных морфологических признаках бактерии отличаются большим разнообразием осуществляемых ими в природе превращений веществ.
Бактерии в совокупности с другими группами микроорганизмов выполняют колоссальную химическую работу. При их участии происходит разложение сложных органических веществ — растительных и животных отстатков — до простых минеральных соединений: углекислоты, аммиака, нитратов, сульфатов и др.,— которые вновь ассимилируются растениями, а затем поступают в организм животного. Таким образом на Земле осуществляется в колоссальном масштабе круговорот жизненно необходимых элементов: углерода, азота, серы, фосфора, железа и др., и бактерии являются важнейшим звеном в этом процессе.
Превращая различные соединения, бактерии получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий чрезвычайно разнообразны.
Одни из бактерий нуждаются в готовых органических веществах — аминокислотах, углеводах, витаминах,— которые должны присутствовать в среде, так как сами не могут их синтезировать. Такие микроорганизмы называются гетеротрофами. Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют исключительно за счет углекислоты. Они называются автотрофами.
По своим потребностям гетеротрофы очень разнообразны: некоторые из них нуждаются в большом наборе аминокислот, витаминов, углеводов и т. д.; другие требуют наличия в среде лишь небольшого числа готовых аминокислот, потребности в витаминах у них могут быть ограничены. Есть и такие формы, которые могут сами синтезировать все вещества: белки, сахара, жиры и т. д., если в среде, где происходит их развитие, присутствует всего одно или несколько простых органических соединений. Такие гетеротрофные организмы ближе стоят к автотрофам.
Каждый организм для поддержания жизни и осуществления процессов, совокупность которых составляет обмен веществ, нуждается в постоянном и непрерывном притоке энергии.
Гетеротрофные микроорганизмы получают энергию при окислении органических веществ кислородом или при сбраживании (без участия кислорода).
Типы окислительных процессов в мире бактерий исключительно разнообразны. Эти микроорганизмы могут окислять любые имеющиеся в природе органические вещества. Если бы в природе существовало какое-либо органическое вещество (продукт животного или растительного происхождения), которое не могло бы быть окислено каким-либо микробом, то оно неизбежно накапливалось бы на поверхности Земли, а этого не происходит. Только в недрах, изолированных от кислорода, могут сохраняться органические вещества — нефть, уголь. Против микробного окисления не могут устоять даже искусственно полученные синтетические вещества, отсутствующие в природе. Но не каждый вид бактерий может разлагать все органические вещества.
Есть формы, приспособленные к использованию лишь небольшого числа веществ, есть и более универсальные.
Более того, бактерии способны окислять не только органические, но и неорганические соединения. Окисление бактериями неорганических веществ — серы, аммиака, нитратов, соединений железа, водорода и др., в процессе которого происходит синтез органических веществ из углекислоты, называется хемосинтезом, а бактерии, осуществляющие этот процесс,— хемосинтетиками.
Различные вещества могут окисляться не только кислородом воздуха, но и соединениями, богатыми кислородом: нитратами, сульфатами и карбонатами. Денитрифицирующие и такие специализированные бактерии, как десульфати-рующие и метановые, в анаэробных условиях могут окислять органические, а также неорганические вещества при помощи этих соединений, которые при этом восстанавливаются соответственно до азота, аммиака, водорода и метана.
Особенностью окисления органических веществ бактериями, как и другими микробами, является то, что оно не обязательно идет до конца как дыхание, т. е. до образования углекислого газа и воды, и в среде остаются продукты неполного окисления.
Механизмы окислительных процессов у микроорганизмов часто включают те или иные стадии дыхания. Огромное разнообразие окисляемых веществ предполагает существование разных механизмов окисления.
Дыхание
Дыхание является самой совершенной формой окислительного процесса и наиболее эффективным способом получения энергии. Главное преимущество дыхания состоит в том, что энергия окисляемого вещества —• субстрата, на котором микроорганизм растет, используется наиболее полно. Поэтому в процессе дыхания перерабатывается гораздо меньше субстрата для получения определенного количества энергии, чем, например, при брожениях.
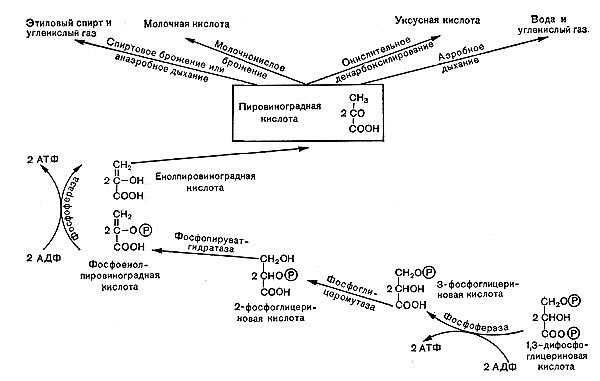
Рис.10. Роль пировнноградной кислоты в процессах дыхания и брожения.
Процесс дыхания заключается в том, что углеводы (или белки, жиры и другие запасные вещества клетки) разлагаются, окисляясь кислородом воздуха, до углекислого газа и воды. Выделяющаяся при этом энергия расходуется на поддержание жизнедеятельности организмов, рост и размножение. Бактерии вследствие ничтожно малых размеров своего тела не могут накапливать значительного количества запасных веществ. Поэтому они используют в основном питательные соединения среды.
В общем виде дыхание можно представить следующим уравнением:
С6Н12О6 + 602 = 6С02 + 6Н20 + 2,87-106 дж.
глюкоза кислород углекислый газ вода энергия
За этой простой формулой скрывается сложная цепь химических реакций, каждая из которых катализируется специфическим ферментом.
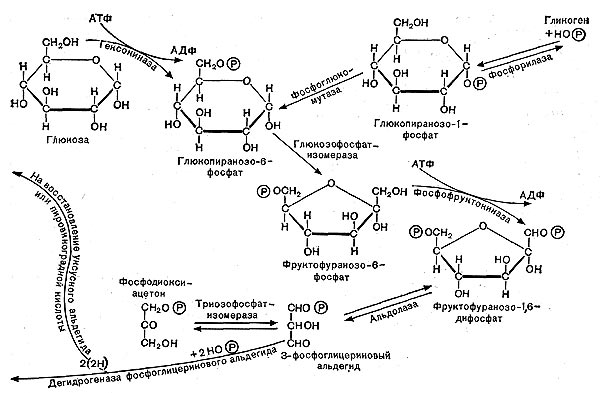
Рис. 11. Схема гликолитического пути расщепления углеводов.
Ферментативные реакции, происходящие в процессе дыхания, в настоящее время хорошо изучены. Схема реакций оказалась универсальной, т. е. в принципе одинаковой у животных, растений и многих микроорганизмов, в том числе бактерий. Процесс дыхания при окислении глюкозы складывается из следующих основных этапов (рис. 10).
Сначала происходит образование фосфорных эфиров глюкозы — моноaктивированная глюкоза в форме дифосфата далее расщепляется на два триозофосфата (трехуглеродные соединения): фосфоглицериновый альдегид и диоксиацетон-фосфат, которые могут обратимо превращаться друг в друга.
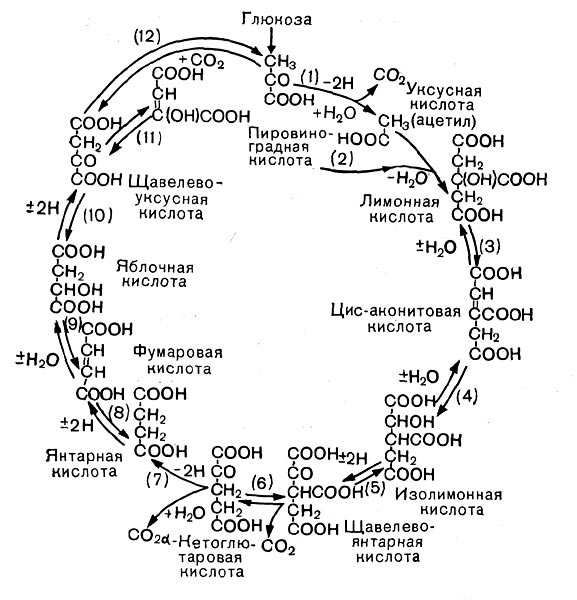
Рис. 12. Цикл трикарбоновых кислот. Стрелками показано направление, а номерами — порядок реакций.
Далее в обмен вступает фосфоглицериновый альдегид, он окисляется в дифосфо-глицериновую кислоту. Назначение этого процесса заключается в отщеплении атомов водорода от окисляемого субстрата и переносе водорода с помощью специфических окислительных ферментов к кислороду воздуха (см. рис. 10, 11).
Водород от фосфоглицеринового альдегида присоединяется к ферменту — никотин-амиддинуклеотиду (НАД); при этом альдегид окисляется до кислоты и выделяется энергия. Часть этой энергии тратится на образование АТФ; при этом присоединяется фосфорная кислота к аденозиндифосфат у— АДФ. При гидролизе АТФ энергия освобождается и может быть затрачена на различные процессы синтеза белка и другие нужды клетки.
Фосфоглицериыовая кислота окисляется до пировиноградной кислоты. При этом также образуется АТФ, т. е. запасается энергия.
На этом завершается первая — анаэробная — стадия процесса дыхания, которая носит название гликолитического пути или пути Эмбдена — Мейергофа — Парнаса. Для осуществления этих реакций кислород не требуется. Образовавшаяся пировиноградная кислота (СН3СОСООН) является интереснейшим и очень важным соединением. Пути расщепления глюкозы в процессе дыхания и многих брожений, вплоть до образования пировиноградной кислоты, идут совершенно одинаково, что впервые было установлено русским биохимиком С. П. Костычевым. Пировиноградная кислота является тем центральным пунктом, от которого расходятся пути дыхания и брожений, откуда начинается специфическая для данного процесса цепь ферментативных превращений -специфическая цепь химических реакций (рис. 11).
В процессе дыхания пировиноградная кислота вступает в цикл трикарбоновых кислот (рис. 12). Это сложный замкнутый круг превращений, в результате которых образуются органические кислоты с 4, 5 и 6 атомами углерода (яблочная, молочная, фумаро-вая, а-кетоглутаровая и лимонная) и отщепля-«тся углекислота.
Прежде всего от пировиноградной кислоты, содержащей три атома углерода, отщепляется СО2 — образуется уксусная кислота, которая с коферментом А образует активное соединение — ацетилкоэнзим А. Он передает остаток уксусной кислоты (ацетил) на щавелевоуксусную кислоту {4 атома углерода), и образуется лимонная кислота (6 атомов углерода). Лимонная кислота претерпевает несколько превращений, в результате выделяется С02 и образуется пяти-углеродное соединение — а-кетоглутаровая кислота. От нее тоже отщепляется С02 (третья молекула углекислого газа), и образуется янтарная кислота (4 атома углерода), которая затем превращается в фумаровую, яблочную и, наконец, щавелевоуксусную кислоту.
На этом цикл замыкается. Щавелевоуксусная кислота снова может вступить в цикл.
Таким образом, в цикл вступает трехуглерод-ная пировиноградная кислота, и по ходу превращений выделяются 3 молекулы С02.
Водород пировиноградной кислоты, освобождающийся при дегидрировании в аэробных условиях, не остается свободным — он поступает в дыхательную цепь (так же, как водород глицеринового альдегида, отнятый при превращении его в глицериновую кислоту). Это — цепь окислительных ферментов.
Ферменты, которые первыми берут на себя водород от окисляемого субстрата, называются первичными дегидрогеназами.
В их состав входят ди- или трипиридин-нуклео-тиды: НАД или НАДФ и специфический белок. Механизм присоединения водорода — один и тот же:
Окисляемое вещество — Н2 + НАД -> окисленное вещество + НАД*Н2
Водород, полученный дегидрогеназой, затем присоединяется к следующей ферментной системе — флавиновым ферментам (ФМН или ФАД).
От флавиновых ферментов электроны попадают на цитохромы — железосодержащие протеиды (сложные белки). По цепи цитохро-мов передается не атом водорода, а только электроны. При этом происходит изменение валентности железа: Fe++ — e->Fe++
Заключительная реакция дыхания — это присоединение протона и электрона к кислороду воздуха и образование воды. Но прежде происходит активирование молекулы кислорода под действием фермента цитохромоксидазы. Активирование сводится к тому, что кислород приобретает отрицательный заряд за счет присоединения электрона окисляемого вещества. К активированному кислороду присоединяется водород (протон), образуя воду.
Кроме упомянутой цепи переносчиков электронов и водорода, известны и другие. Процесс этот гораздо более сложен, чем изложенная схема.
Биологический смысл этих превращений заключается в окислении веществ и образовании энергии, В результате окисления молекулы сахара (глюкозы) в АТФ запасается 12,6-1053ж энергии, в самой молекуле сахара содержится 28,6-106 дж, следовательно, полезно используется 44% энергии. Это очень высокий коэффициент полезного действия, если сравнить его с к. п. д. современных машин.
В процессе дыхания образуется огромное количество энергии. Если вся она выделилась бы сразу, то клетка перестала бы существовать. Но этого не происходит, потому что энергия выделяется не вся сразу, а ступенчато, небольшими порциями. Выделение энергии небольшими дозами обусловлено тем, что дыхание представляет собой многоступенчатый процесс, на отдельных этапах которого образуются различные промежуточные продукты (с разной длиной углеродной цепочки) и выделяется энергия. Выделяющаяся энергия не расходуется в виде тепла, а запасается в универсальном макроэр-гическом соединении — АТФ. При расщеплении АТФ энергия может использоваться в любых процессах, необходимых для поддержания жизнедеятельности организма: на синтез различных органических веществ, механическую работу, поддержание осмотического давления протоплазмы и т. д.
Дыхание является процессом, дающим энергию, однако его биологическое значение этим не ограничивается. В результате химических реакций, сопровождающих дыхание, образуется большое количество промежуточных соединений. Из этих соединений, имеющих различное количество углеродных атомов, могут синтезироваться самые разнообразные вещества клетки: аминокислоты, жирные кислоты, жиры, белки, витамины.
Поэтому обмен углеводов определяет остальные обмены веществ (белков, жиров). В этом его огромное значение.
С процессом дыхания, его химическими реакциями связано одно из удивительных свойств микробов — способность испускать видимый свет — люминесцировать.
Известно, что ряд живых организмов, в том числе бактерии, могут испускать видимый свет. Люминесценция, вызываемая микроорганизмами, известна уже в течение столетий. Скопление люминесцирующих бактерий, находящихся в симбиозе с мелкими морскими животными, иногда приводит к свечению моря; с люминесценцией встречались также при росте некоторых бактерий на мясе и т. д.
К основным компонентам, взаимодействие между которыми приводит к испусканию света, относятся восстановленные формы ФМН или НАД, молекулярный кислород, фермент люцифераза и окисляемое соединение — люциферин. Предполагается, что восстановленные НАД или ФМН реагируют с люциферазой, кислородом и люциферином, в результате чего электроны в некоторых молекулах переходят в возбужденное состояние и возвращение этих электронов на основной уровень сопровождается испусканием света. Люминесценцию у микробов рассматривают как «расточительный процесс», так как при этом энергетическая эффективность дыхания снижается.
Брожение
Жизнь микробов возможна и без доступа кислорода воздуха. Энергия, необходимая для жизнедеятельности организма, в этих условиях образуется в результате процессов брожения. Наиболее распространены виды брожений, в процессе которых происходит распад органических веществ (преимущественно Сахаров) под влиянием микроорганизмов, представляющий совокупность окислительно-восстановительных реакций. Брожения никогда не приводят к полному окислению органических веществ. Многие характерные формы брожения протекают без участия кислорода воздуха — анаэробно.
Поскольку свободный кислород, имеющийся на нашей планете, образовался в результате фотосинтеза, возникшего на более поздних этапах развития жизни на Земле, совершенно очевидно, что анаэробный способ извлечения энергии — брожение — более древний, чем процесс дыхания.
Брожение известно людям с незапамятных времен. Тысячелетиями человек пользовался спиртовым брожением при изготовлении вина. Еще раньше было известно о молочнокислом брожении. Люди употребляли в пищу молочные продукты, готовили сыры. При этом они не подозревали, что эти процессы происходят с помощью микроорганизмов. Термин «брожение» был введен голландским алхимиком Ван Хельмонтом в XVII в. для процессов, идущих с выделением газов (fermentatio — кипение). Затем в XIX в. основоположник современной микробиологии Луи Пастер показал, что брожение является результатом жизнедеятельности микробов, и установил, что различные брожения вызываются разными микроорганизмами.
Спиртовое брожение
Спиртовое брожение — это процесс окисления углеводов, в результате которого образуются этиловый спирт, углекислота и выделяется энергия.
Сбраживание cахаров известно с глубокой древности. В течение столетий пивовары и виноделы использовали способность некоторых дрожжей вызывать спиртовое брожение, в результате которого сахара превращаются в спирт.
Брожение производят главным образом дрожжи, а также некоторые бактерии и грибы. В различных странах для получения спирта используют различные микроорганизмы. Например, в Европе используют в основном дрожжи из рода Saccharomyces, в Южной Америке — бактерии Pseudomonas lindneri, в Азии — мукоровые грибы.
Сбраживаться могут лишь углеводы, и притом весьма избирательно. Дрожжи сбраживают только некоторые 6-углеродные сахара (глюкозу, фруктозу, маннозу).
Схематично спиртовое брожение может быть изображено уравнением
С6Н12О6 -> 2С2Н5ОН + 2С02 + 23,5 • 104 дж
глюкоза этиловый спирт углекислота энергия
Процесс спиртового брожения — многоступенчатый, состоящий из цепи химических реакций. Превращения глюкозы до образования пи-ровиноградной кислоты происходят так же, как и при дыхании. Эти реакции происходят без участия кислорода (анаэробно). Далее пути дыхания и брожения расходятся.
При спиртовом брожении пировиноградная кислота превращается в конечном итоге в спирт и углекислоту. Эти реакции протекают в две стадии. Сначала от пирувата отщепляется С02 и образуется уксусный альдегид; затем уксусный альдегид присоединяет водород, восстанавливаясь в этиловый спирт. Все реакции катализируются ферментами. В восстановлении альдегида участвует НАД-H2.
Обычно при спиртовом брожении, кроме главных продуктов, образуются побочные. Они довольно разнообразны, но присутствуют в небольшом количестве: амиловый, бутиловый и другие спирты, смесь которых называется сивушным маслом — соединение, от которого зависит специфический аромат вина. Образование побочных веществ связано с тем, что превращение глюкозы частично идет другими путями.
Биологический смысл спиртового брожения заключается в том, что образуется определенное количество энергии, которая запасается в форме АТФ, а затем расходуется на все жизненно необходимые процессы клетки.
Дата добавления: 2020-11-18; просмотров: 657;
Поиск по сайту
Узнать еще
- I. 3. Функции минеральных веществ плазмы крови
- III. Биогеохимические циклы элементов и веществ (на примере основных биогенных элементов: углерод, азот, фосфор) и их количественные характеристики.
- III. В зависимости от цели обмена, результатов той или иной деятельности различают коммерческий и некоммерческий маркетинг.
- III. Характеристики основных классов загрязняющих веществ в водной среде.
- IV. Основные источники поступления загрязняющих веществ в водную среду.
- VIII. 2. Большой круговорот вещества и роль в нем человека
- Абиотическая миграция вещества
- Аварии с выбросом (угроза выброса) радиоактивных веществ (РВ)

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
