Центральные хеморецепторы
Центральные хеморецепторы обнаружены в продолговатом мозге на вентромедиальной поверхности на глубине не более 0.2 мм. В этой области расположены два рецептивных поля (рисунок 15), обозначаемые буквами M и L, между ними обнаружено небольшое поле S. Поле S не чувствительно к химизму среды, но его разрушение приводит к исчезновению эффектов возбуждения полей M и L, Этой промежуточной зоне принадлежит важная роль в передаче информации от полей M и L непосредственно дыхательным вентральным и дорзальным ядрам, и передаче информации ядрам другой стороны продолговатого мозга.
В этой же области проходят аферентные пути от периферических хеморецепторов. В вентролатеральных отделах, в районе хеморецептивных полей расположены структуры, оказывающие существенное влияние на тонус вегетативной нервной системы. Вероятно, эта зона имеет отношение к интеграции ритма дыхания и легочной вентиляции с системой кровообращения. В частности в зонах S и М есть нейроны, которые имеют связи с грудными сегментами спинного мозга, их раздражение приводит к повышению сосудистого тонуса. Часть нейронов этой области активируется при раздражении аортального и синокаротидного нервов (информация от периферических хемо- и барорецепторов каротидного синуса и дуги аорты), часть нейронов отвечают на раздражение ядер гипоталамуса (информация об осмотической концентрации внутренней среды, температуре). Таким образом, структуры S и М полей интегрируют афферентные сигналы от расположенных выше нейронных образований и передают тонизирующие влияния вазоконстрикторным нейронам спинного мозга. Каудальный отдел, поле L демонстрирует при его электрическом раздражении противоположные эффекты. Вместе с тем существует четкая нейронная обособленность между нейронами, регулирующими функции кровообращения и нейронами, связанными с дыхательным центром.
Рисунок 15. Расположение хеморецепторов на вентральной поверхности продолговатого мозга
M, L, S поля, участвующие в хеморецепции.
Р – мост,
П – пирамида,
V и XII – черепномозговые нервы,
С1 первый спинномозговой корешок
В настоящее время совершенно точно установлено, что центральные хеморецептивные нейроны возбуждаются только при действии на них ионов водорода. Каким же образом повышение напряжения СО2 приводит к возбуждению этих структур? Оказывается хемочувствительные нейроны расположены во внеклеточной жидкости и воспринимают изменения рН, вызванные динамикой СО2 в крови.
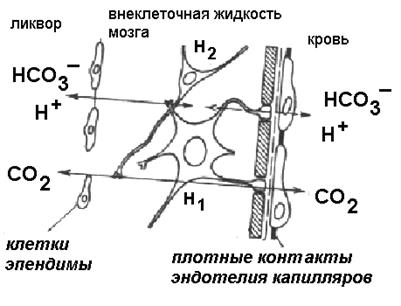
Вентролатеральные отделы продолговатого мозга представлены нервными клетками, астроцитарной глией, развитой мягкой мозговой оболочкой и окружены тремя средами мозга: кровью, ликвором и внеклеточной жидкостью (рисунок 16). Среди нейронов выявляются крупные мультиполярные клетки и мелкие, округлые. Оба типа нейронов образуют небольшое ядро, которое контактирует с прилежащими ядрами ретикулярной формации. Крупные мультиполярные нейроны имеют периваскулярную локализацию и их отростки располагаются вблизи стенок микрососудов. В механизме хеморецепции в настоящее время остается много непонятного. Перечислим факты, которые установлены и помогают объяснить этот механизм
Мультиполярные нейроны всегда увеличивают свою метаболическую и электрическую активности при гиперкапнии и при локальном повышении концентрации ионов водорода во внеклеточной жидкости, омывающей эти нейроны.
Между напряжением СО2 в альвеолярном воздухе и в артериальной крови, с одной стороны, и рН внеклеточной жидкости мозга с другой стороны существует линейная зависимость.
И гиперкапния, и локальное повышение рН внеклеточной жидкости всегда сопровождаются дыхательной реакцией - увеличение глубины и частоты дыхания.
Между ликвором и кровью существует незначительная, но устойчивая разность потенциалов.
Снижение рН приводит к изменению этой разности потенциалов.
Существует градиент концентрации по ионам водорода между кровью и внеклеточной жидкостью - во внеклеточной жидкости ионов водорода больше. Градиент поддерживается активным переносом протонов из крови во внеклеточную жидкость.
На границе между кровью и внеклеточной жидкостью высока активность фермента карбоангидразы.
Эндотелий сосудов, граничащий с внеклеточной жидкостью в области хеморецептивных полей не проницаем для ионов Н+ и НСО3- но хорошо проницаем для СО2 .
Приблизительно схему событий можно представить следующим образом: 1) повышение концентрации СО2 в крови и его свободная диффузия через зону с высокой карбоангидразной активностью 2) СО2 соединяется с Н2О под влиянием карбоангидразы, затем диссоциирует с освобождением Н+. 3) накопление во внеклеточной жидкости ионов водорода приводит к повышению активности мультиполярных нейронов.
 Одновременно происходит уменьшение разности потенциалов между кровью и ликвором. Эти события служат мощным афферентным стимулом для дыхательного центра. Следует обратить внимание на высокую чувствительность всех структур к изменению рН - изменение потенциала и дыхательная реакция отмечаются при снижении рН крови на 0.01 единицу. Высока и надежность этих структур - мультиполярные нейроны способны изменять свою активность в диапазоне рН от 7 до 7,8 , такие изменение в норме невозможны.
Одновременно происходит уменьшение разности потенциалов между кровью и ликвором. Эти события служат мощным афферентным стимулом для дыхательного центра. Следует обратить внимание на высокую чувствительность всех структур к изменению рН - изменение потенциала и дыхательная реакция отмечаются при снижении рН крови на 0.01 единицу. Высока и надежность этих структур - мультиполярные нейроны способны изменять свою активность в диапазоне рН от 7 до 7,8 , такие изменение в норме невозможны.
Рисунок 16 Локализация мультиполярных нейронов (хемосенсоров) относительно внутренних сред мозга: крови, внеклеточной жидкости мозга и ликвора.
Н1 – крупный мультиполярный нейрон, Н2 мелкий мультиполярный нейрон,
Итак, важнейшим физиологическим свойством центрального хеморецептивного механизма является изменение активности нейронов в прямой зависимости от концентрации ионов водорода во внеклеточной жидкости мозга. Основная задача этого механизма - информировать дыхательный центр об отклонения рН, а следовательно, и концентрации СО2 в крови.Обратите внимание на то, что саморегулирование в этом случае будет осуществляться по принципу отклонения от физиологической нормы.
Дата добавления: 2020-11-18; просмотров: 721;
Поиск по сайту
Узнать еще
- Артериальные хеморецепторы
- Высшие (центральные) органы государственной власти КНР
- Глава II. РЕФЛЕКТОРНО-ДВИГАТЕЛЬНАЯ ФУНКЦИЯ, ПЕРИФЕРИЧЕСКИЕ И ЦЕНТРАЛЬНЫЕ ПАРАЛИЧИ
- Главные центральные моменты инерции сложных сечений произвольной формы
- Главные центральные моменты инерции сложных симметричных сечений
- Органы иммунной системы – центральные и периферические. Лимфатическая система, ее взаимоотношения с иммунной системой.
- Расчет координат пикетов через центральные углы
- Структура системы федеральных государственных органов исполнительной власти (высшие, центральные, территориальные органы государственного управления)

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
