ПАЙДА БОЛУ МЕХАНИЗМІ. 2 глава
Керісінше жасаушадағы су молекулалары толығымен сыртқа шықса, яғни ағзаның сусыздануы байқалса, онда жасуша жиырлып, оның жансыздануы орын алады, яғни жасуша өмір сүруін тоқтатады. Мұндай құбылыс «коллапс» деп аталынады. Қан тамарлар жүйесіндегі қанның осмостық қысымын тұрақты деңгейде реттеп отыратын арнаулы альбумин деген ақуыз бар. Ағзадағы коллапс құбылысы көп қан кетудің әсерінен емес, қанның осмостық қысымының бірден төмендеуінен байқалады екен, сондықтан көп қан жоғалту кезінде ағзаға инерті жоғарымолекулалы қан ауыстырғыштарды салады, нәтижесінде қанның осмостық қысымы өз деңгейіне келеді.
5. Фильтация (сүзу,сүзгі) деп гидростатикалық қысым градиенті есебінен су молекулаларының мембрана порлары арқылы тасымалдануын атайды. Су молекулаларының тасымалдану жылдамдығы Пуайзель заңы бойынша жүреді: dV/dt = P1 – P2/w, мұндағы dV/dt- суды тасымалдау жылдамдығы, w- гидравликлық қысым, ол w = 8lh/pr4 тең, l - пор ұзындығы, r- оның радиусы, h- судың тұтқырлдық коэффициенті. Фильтрация құбылысы қан тамырлары арқылы су молекуласын тасымадануда маңызды орын алады, кейбір патологияларда фильтрация күшейіп нәтижесінде дене ісінеді.
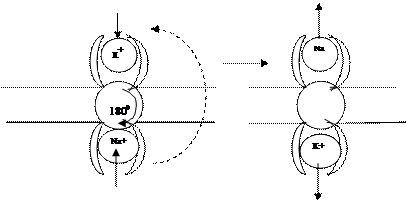
Активті тасымалдау(АТ). Егер мембранада тасмалдау тек пассивті түрде жүретін болса, онда мембрананың ішкі және сырты ортадағы иондар концентрациясы теңесер еді, бұл жасуша үшін өте қауіпті жағдай, сондықтан мембрана орталарындағы иондардың концентрацияларын әр түрлі болуын қамтамасыз ететін механизм де болуы тиіс. Ол активті тасымалдау нәтижесінде іске асады және заттар концентрациясы аз ортадан концентрациясы көп ортаға қарай, яғни градиентке қарсы бағытта тасымалданады, әрине мұндай тасымалдануға энергия қажет. Осы мақсаттағы энергия көзі болып аденозин трифосфат қышқылы молекуласының (АТФ) ыдырау кезінде бөлінетін энергиясы қолданылады (5 сурет).
 3 Na+
3 Na+
2 K+
3 Na+ 2 K+
5 сурет
Ғылыми тәжірибе негізінде АТФ бір молекуласы ыдырағанда бөлінетін энергия арқылы сыртқы ортаға 3 натрии ионын, ішкі ортаға 2 калии ионын тасымалдауға жететіндігін көрсетті. Тасымалдағыш ақуыз АТФ молекуласынан бөлінген энергияның есебінен бір жағымен сыртқы ортадан 2 калии ионын, ішкі ортадан 3 натрии ионын қосып алып, мембрана қабатында 1800 бұрылып, натрии ионын сыртқы ортаға, калии ионын ішкі ортаға жеткізеді, онан соң ақуыз қайта өз орнына келеді.
АТФ энергиясы арқылы зат тасымалдайтын осындай ақуыздарды иондық насостар деп атайды. Қазіргі кезде толық зерттелген осындай үш түрлі электрогенды насостар белгілі, олар: калии-натрии насосы (3 натрии ионын сыртқа, 2 кали ионын ішке), кальции насосы (2 кальци ионын сыртқа) және протон насосы (2 протонды сыртқа)
Осындай тасымалдау арқылы жасуша ішкі ортада калий ионының концентрациясын жоғары деңгейде, ал натрии ионынын төмен деңгейде ұстап тұрады. Активті тасымалдау кезінде мембрананың талғампаздық (селективті) қасиеті сақталады.
Заттар мембранадағы иондық каналдар арқылы да тасымалданады. Мембрана қабатындағы орналасқан ақуыз молекулалары мен липидтер саңлаулар (пор), яғни иондық каналдар жасайды. Мұндай индық каналдар арқылы мембранаға су молекулалары мен ірі иондар тасымалданады. Каналдар мембрананың өткізгіштік коэффициентін жоғарылатады, канал көп және олардың радиустары үлкен болған сайын мембрананың өткізгіштігі де артады. Каналдардың тасымалданатын заттарды таңдап өткізетін қасиеті бар. Әр иондық канал өзіне тиісті ғана ионды немесе затты өткізеді, яғни натрии каналы негізінен натрий ионын, калий каналы тек калии иондарын өткізеді. Сонымен қатар каналдардың заттарды өткізуі олардың зарядына байланысты болады, мысалы катиондарды өткізетін канал аниондарды өткізбейді, керісінше аниондарды өткізетін канал катионды өткізбейді. Каналдар өзіне тән емес иондарды да өткізеді, бірақ ол заттар үшін каналдың өткізгіштігі өте төмен, мысалы натрии каналының калии ионын өткізуі натримен салыстырғанда 20 есе төмен.
Мембранадағы каналдарды кейбір қосылыстар арқылы жауап тастауға болады екен, мысалы тетродотоксин молекуласы натрии каналын, тетраэтиламония молекуласымен калии каналын жауап тастайды. Мұндай әдістер медицинада көп қолданылады, мысалы ота жасағанда, тіс жұлғанда қажетті аймақты жансыздандыру осы құбылыстардың арқасында орындалады.
1842 жылы Э.В.Брюкке қуық қалтасындағы суды тасымаладауды зерттеу нәтижесінде тірі ағза ұлпаларында зат тасымалы саңлаулар (пор) арқылы жүретіндігін жобалаған болатын. Брюкктің бұл гипотезасы тек жүз жылдан кейін Ходжикин мен Хакслидің (1952), Девсон мен Даниеллидің (1955) ғылыми жұмыстары және Хладни мен Хайдонның(1970, 1972) жасанды липид биқабатына орнатылған грамицидитті(ақуыз) каналдың қасиетін зерттеулері нәтижесінде дәлелденді.
Қазіргі заманғы концепция бойынша мембранадағы иондық каналдар деп липид биқабатын тесіп орналасқан, электрохимиялық потенциалы аз жаққа қарай зат тасымалын қамтамасыз ететін интегралды ақуыздарды (олардың комплексін немесе гликопротеид) атайды. Иондық канал арқылы зат тасымылының өткізгіштік коэффициенті 10-8 – 10-9 м/с тең, бірақ бұл шама судағы еркін диффузия арқылы иондарды тасымалдаудан 5-6 есе төмен.
Хилле 1977-84 ж.ж. аралығында жүргізген зерттеулері нәтижесінде иондық каналдың мынадай моделін ұсынды: иондық каналда селективті сүзгі (фильтр) және «қақпа» тәрізді механизм бар. (6- сурет).
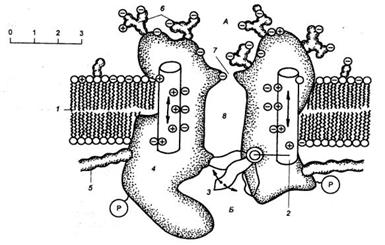
6 сурет. 1 - липидті биқабат, 2- кернеу сенсоры, 3- қақпа, 4 - селективті фильтр 5- ақуыз макромолекуласы, 6-көмір сулар, 7- пор,
Селективті сүзгі канал арқылы тек белгілі бір заттарды өткізу немесе оларды басқалардан бөліп алумен айналысады. Бірақ мұндай ион каналының өлшемі (радиусы, кеңдігі) өзгермейді, мембрананың өткізгішті әр уақытта тұрақты, өзгеріссіз қалады.
Мембараның өткізгіштігі «қақпа» механизмі арқылы реттеледі. Иондық каналдың «қақпасы» ақуыз комплексінен құралады, «қақпаның сыртқы механохимиялық және т.б. әсерлерден «бұралатын» немесе «кері бұралатын» қасиеті бар, соның нәтижесінде ақуыз комплексінің саңлауы тарылады немесе кеңейеді, яғни «қақапының» өткізгіштігі өзгеріп тұрады.
Ақуыз комплексіндегі канал саңлауының өлшемі тасымалданатын зат өлшемімен шамалас болып келеді. Канал жабық не толық ашық күйде болады және ол арқылы зат бір жақты бағытта тасымалданады, соңғы күйде зат тасымалдау қарқыны максимал деңгейінде жүреді. Мысалы, натрии және калий иондарын тасмалдау жылдамдығы 107 ион/с тең.
Мембрананың нақты бір зат үшін өткізгіштігі берілген уақыт мезгіліндегі ашық тұрған ион каналдарының санын тәуелді екен. Мембрананың өткізгіштігі мына өрнекпен сипатталады: p= npr2D/l, мұндағы n- мембрананың бірлік ауданына сәйкес келетін ашық канал саны, r – канал радиусы, D- заттың судағы дифузия коэффициенті, l- каналдың ұзындығы( ол мембаран қалыңдығығна тең).
Иондық каналдардың «қақпаларының» ашық, не жабық күйде болуын реттейтін бірнеше факторлар бар. Егер канал «қақпасының» күйі мембранадағы потенциалдың өзгеруіне байланысты болса оны потенциалға тәуелді каналдар деп атайды. Мұндай каналдағы «қақпаның» күйі мембранадағы кернеу өзгерісін сезгіш жүйе (сенсор) арқылы реттеледі. Сенсор канал құрылымына енетін ақуыз комплекісін құрайтын аминоқышқыл қалдықтарынан құралған байланысқан зарядтардан тұрады. Мысалы, потенциалға тәуелді натрий каналында кернеу сезгіш жүйе - сенсор аминоқышқыл қалдықтарынының (гистидилом,лизилом, аргинилом және т.б.) катиондарынан құрылған.
Иондық каналдардың екінші түрі потенциалға тәуелсіз болып келеді. Мұндай каналдардың өткізгіштігі мембранадағы потенциалдың өзгерісінен емес, керісінше басқа түрдегі, мысалы, химиялық, механикалық, сәулелік және т.б. әсерлерден туындайды. Потенциалға тәуелсіз каналдардың «қақпаларының» күйі сыртқы әсерлерді қабылдайтын мембрандағы түрлі рецепторлардан келетін «ақпаратарға» байланысты болады. Рецепторлардан келетін ақпарат жасуша аралық сигналдар жүйесі арқылы мембранадағы «қақпаға» жеткізіледі, ол өз кезегінде ақпарат мәліметіне байланысты «қақпаны» ашады немесе жабады. Иондық каналдардың мұнан да басқа түрде жұмыс істейтіндері бар екендігі анықталған.
Иондық каналдардың тағы бір маңызды қасиеті ионның түрі мен оның химиялық құрылымын ажырата алуы. Иондық канал талғампаздығының табиғатын білу жасушаның электрлік белсенділігінің молекулалық механизімін анықтауға мүмкіндік береді. Иондық талғампаздық туралы бірнеше гипотезе бар.
ТАҚЫРЫП БОЙЫНША СТУДЕНТТІҢ ӨЗІ ӨЗІ ТЕКСЕРУІНЕ АРНАЛҒАН ТЕСТ СҰРАҚТАРЫ
1. Биологиялық мембранадағы липитдтер биомолекулалық қабат түрiнде орналасқан деген тұжырымның дұрыстығы оның ... өлшеу барысанда дәлелдендi.
A. |электрлiк параметрлерiн
B. |температурасын
C. |қысымын
D. |қалыңдығын
E. |тығыздығын
2. Жасуша ашық термодинамикалық жүйе болғандықтан ол өзiн қоршаған ортамен ...
A. |үнемi энергия, зат және ақпарат алмасады.
B. |кейде энергия алмасады.
C. |әсерлеспейдi.
D. |анда-санда жылу алмасады.
E. |электрон алмасады.
3. Диффузия деп заттардың ... тасымалдануын айтады.
A. |өз еркiмен концентрациясы көп ортадан аз ортаға қарай
B. |ерiксiз концентрациясы көп ортадан аз ортаға қарай
C. |өз еркiмен концентрациясы аз ортадан көп ортаға қарай
D. |ерiксiз концентрациясы аз ортадан көп ортаға қарай
E. |энергия әсерiнен
4. Диффузия жылдамдығы ... анықталынады.
A. |иондардың қозғалғыштығымен
B. |иондардың зарядымен
C. |ортаның температурасымен
D. |иондардың мөлшерiмен
E. |жасушаның күйiмен
5. Биомембрана қабабты арқылы диффузия құбылысы ... жүредi.
A. |концентрациялық градиент есебiнен
B. |электрлiк градиент есебiнен
C. |өз еркiмен концентрациясы аз ортадан көп ортаға қарай
D. |осмостық градиент есебiнен
E. |энергия есебiнен
6 Жеңiлдетiлген диффузия ... жүредi.
A. |тасымалдағыш молекулалардың қатысуымен
B. |электрондардың әсерiнен
C. |қышқылдық заттардың қатысуымен
D. |температураның өзгеруiнен
E. |сыртқы күштiң әсерiнен
7 Сүзгiштiк (фильтрация) деп ... есебiнен мембрана қуыстары (порлары) арқылы тасымалдануын айтады.
A. |су молекулаларының қысым градиентi
B. |ақуыз молекулаларының қысым градиентi
C. |су молекулаларының концентрациялық градиент
D. |иондардың температурасының өзгеру
E. |иондардың концентрациясы
8 Ағзадағы судың қан тамырлары қабырғалары арқылы өтуiнде сүзгiштiк құбылысының атқаратын рөлi үлкен, соның арқасында кейбiр патологиялық өзгерiс кезiнде сүзгiштiк ...
A. |ұлғаяды, соның себеiнен дене iсiнедi.
B. |кемидi, соның себеiнен дене iсiнедi.
C. |ұлғаяды, соның себеiнен дене кiшiрейiп, семедi.
D. |ұлғаяды, ал денеде өзгерiс байқалмайды.
E. |өзгермейдi.
9 Осмос деп жартылай өткiзгiш мембрана арқылы (суда ерiген заттар үшiн өтiмсiз, су үшiн өтiмдi) ... тасымалдағыш молекулалардың қатысуымен тасымалдануын айтады.
A. |ерiген заттардың концентрациясы көп ортадан аз ортаға қарай су молекулаларының
B. |ерiген заттардың концентрациясы аз ортадан көп ортаға қарай су молекулаларының
C. |қышқылдық заттардың
D. |электрондардың
E. |сыртқы күштiң әсерiнен
10 Осмос деп - ... тасымалдануын айтады.
A. |жартылай өткiзгiштi мембрана арқылы су молекулаларының концентрациясы көп ортадан аз ортаға
B. |қысым градиентi әсерiнен су молекулаларының
C. |заттардың өз еркiмен концентрациясы көп ортадан аз ортаға
D. |заттардың өз еркiмен оптикалық тығыздығы көп ортадан аз ортаға
E. |заттардың өз еркiмен оптикалық тығыздығы аз ортадан көп ортаға
11Фильтрация деп - .... тасымалдануын айтады.
A. |қысым градиентi әсерiнен су молекулаларының
B. |жартылай өткiзгiштi мембрана арқылы су молекулаларының
концентрациясы көп ортадан аз ортаға
C. |заттардың өз еркiмен концентрациясы көп ортадан аз ортаға
D. |заттардың өз еркiмен оптикалық тығыздығы көп ортадан аз ортаға
E. |заттардың өз еркiмен оптикалық тығыздығы аз ортадан көп ортаға
12 Пассивті тасымалдау деп..... тасымалдауды айтады.
A. |заттарды концентрациясы аз ортадан көп ортаға қарай
B. |электрлiк потенциалы көп ортадан аз ортаға қарай
C. |заттарды концентрациясы көп ортадан аз ортаға қарай
D. |температураның өзгеруiнен болатын
E. |сыртқы күштiң әсерiнен
13 Активті тасымалдау деп ... тасымалдауды айтады.
A. |заттарды концентрациясы аз ортадан көп ортаға қарай
B. |электрлiк потенциалы көп ортадан аз ортаға қарай
C. |заттарды концентрациясы көп ортадан аз ортаға қарай
D. |температураның өзгеруiнен болатын
E. |сыртқы күштiң әсерiнен
14 Фик теңдеуі
A. |J = D dC/dx
B. | J = -D dC/dx
C. | J = -D dX/dC
D. | J = D dХ/dC
E. J = D/ dC×dx
15. Нернст –Планк теңдеуі
A. |J = D dC/dx + cbZFdj/dx
B. | J = -D dC/dx+ cbZFdj/dx
C. | J = -D dX/dC + cbZFdj/dx
D. | J = D dХ/dC + cbZFdj/dx
E. J = D/ dC×dx + cbZFdj/dx
3 дәріс. БИОЭЛЕКТРЛІК ПОТЕНЦИАЛ
Лекция жоспары
1. Потенциал туралы түсінік
2. Биопотенциал түрлері: тыныштық және әсер потенциалдары
3. Гольдман-Ходжкин-Катц теңдеуі.
4. Әрекет потенциалының пайда болу механизмі.
5. Ходжкин-Хаксли теңдеуі.
6. Әрекет потенциалының таралуы.
Лекция мақсаты: биопотенциалдың түрлерін, пайда болу механизімін, оның математикалық өрнегін және әрекет потенциалының нерв талшығымен таралуын талдау.
Физикада потенциал термині әр түрлі өрістердің (электростатикалық, магниттік, гравитатциялық т.б.) энергиясын сипаттау үшін қолданылатын шама. Биологиялық мембранада электр өрісі бар, олай болса ол өрістің энергиясын да осы тәрізді бипотенциал деген шамамен сипаттаймыз. Көбіне потенциалдың шамасын қолданбайды, олардың айырымын, яғни екі нүктедегі потенциалдар айрымын қолданады: Dj =j2 - j1, мұндағы j2 и j1 сәйкес нүктелердегі (аймақтардағы) потенциал шамасы. Биопотенциал деп ағзаның екі нүке арасындағы потенциалдар айырымын немесе мембранадағы биопотенциал деп, мембрананың ішкі және сыртқы орта арасындағы потенциалдар айырымын атайды: DjМ =jІШ - jСЫРТ.
Барлық уақытта қозбаған жасушада потенциал айырымы кездеседі, оны тыныштық потенциалы деп атайды, жасуша ішінде потенциал «теріс», сыртында «оң» таңбалы болады.
Биопотенциалдың пайда болу механизімін толық талдау үшін, біз алдымен олардың негізінде қандай физика-химиялық құбылыстардың жатқандығын, және потенциалдың шамасы неге байланысты болатындығын талдайық.
Пайда болу механизміне байланысты потенциалдар: диффузиялық, фазалық және мембраналық деген түрлерге бөлінеді. Диффузиялық потенциал сұйық ортаны бөліп тұрған шекарадағы иондардың қозғалғыштығының әр болуынан пайда болады (1 сурет).
HCl Н2О 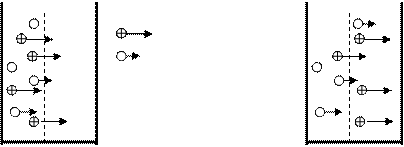 j- j+
j- j+
Н+
Cl-
1 cурет
Мысалы, ортасында поры бар бөгет арқылы екіге бөлінген ыдыстың сол жағына тұз қышқылы ертіндісін (HCl), оң жағына су құяйық. Пор тек иондарды өткізсін делік. Диффузия құбылысы әсерінен және екі ортада концентрациялық градиенттің болуы себебінен иондар ыдыстың екінші жағына қарай қозғалады. Концентрациялық градиент деп белгілі бір бағыттағы бірлік ұзындыққа сәйкес келетін концентрация айрымына тең шаманы атайды. Мысалы, ертіндінің бірінші нүктедегі концентрациясы С1, одан l қашықтықта жатқан екінші нүктедегі концентрациясы С2 болса, онда gradC=С1- С2/l тең болады.
Сонымен концентрациялық градиенттің нолден өзгеше болуы және оң зарядталған сутегі иондарының қозғалғыштығының жоғары болуы себепті, оң зарядталған сутегі иондары теріс зарядталған хлор иондарын артқа тастап, ыдыстың оң жағына бұрын жетеді, нәтижесінде ыдыстың оң жақ бөлігі оң зарядталады, ал сол жақ бөлігінде оң зарядтардың аз болуы (сутегі иондарының біраз бөлігі оң жаққа ауысып кетті) теріс зарядтардың басым болуы себепті ол орта теріс зарядталады. Осылайша ыдыстың екі жағындағы екі түрлі зарядтардың арасында электр өрісінің айырымы туындайды оны диффузиялық потенциал деп атаймыз және оның шамасы Гендерсон теңдеуімен анықталынады:
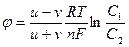
мұндағы u-катионның жылдамдағы, v-анионның жылдамдығы, R- газ тұрақтысы, T- абсолюттік температура, n- ионның валенттілігі, F- Фарадей саны, c1 – бірінші ортадағы ионның концентрациясы, c2 – екініш ортадағы ионның концентрациясы. Бұл өрнектен, дифффузиялық потенциалдың шамасы катион мен анионның қозғалғыштығының айрымашылығына, олардың концентрациясына тікелей байланысты екендігі көрінеді. Егер катион мен анионның жылдамдығы тең болса, немесе олардың концентрациясы бірде болса, онда диффузиялық потенциал нолге тең болады.
Биологиялық жүйелерде потенциалдың бүл түрі негізінен жасушаның механикалық зақымдалуы кезінде байқалады және иондардың зақымдалған аймақтан зақымдалмаған аймаққа қарай диффузиясы жүреді.
Қарастырылған мысалдағы бөгет аниондарды тіптен өткізбесін делік, сонда ыдыстың оң жағына тек сутегі иондары өтеді, яғни v=0 болғандықтан
Гендерсон теңдеуі Нернст теңдеуіне айналады: j= RT/nF ln C1/C2.
Фазалық потенциалдар араласпайтын екі фазалық күйдегі заттар шекарасында (судағы электролит ертіндісі мен май) пайда болады. Мысалы, анионға салыстырғанда катиондар майлы ортада жақсы еритін болса, онда ол майлы ортадаға жылдам ауысып, ортаны оң зарядтайды.
Мембраналық потециалдың пайда болуы туралы алғашқы теорияны 1902 жылы Бернштейн ұсынды. ХХ ғасырдың 50 жылдары Ходжикин, Хакси және Катц, Бернштейнннің теориясының дұрыстығын тәжірибе арқылы дәлелдеді. Олар мембрананың иондарды таңдап өткізуінен мембрананың іші мен сыртқы орталарындағы иондардың концентрациясының әр түрлі болуы биопотенциалдың пайда болуының басты себебі деген қорытынды жасады және оны кальмардың аксонында жүргізген тәжрибелері арқылы толық дәлеледі. Тәжірибе қозбаған мембрананың (кальмар аксоны) калий иондарын жақсы, ал натрии және басқа иондарды нашар өткізетіндігін көрсетті. Оның үстіне калии иондарының концентрациясы жасуша ішінде сыртқы ортамен салыстырғанда 20-40 есе көп, олай болса, диффузия әсерінен калии иондары сыртқа қарай тасымалданады, ал сыртқы ортадағы иондар ішке ене алмайды, олар үшін мембрана өтімділігі өте төмен. Сонымен сыртқы ортаға өткен калии иондары ортаны оң зарядтайды, ішкі ортадаға оң зарядтардың біраз бөлігін калий алып кеткендіктен, ішкі орта теріс зарядты болады (ішкі ортадағы теріс зарядтар сыртқа өте алмайды, оң зарядтар азайды). Бұл үрдіс динамикалық тепе теңдік орнағанға дейін жүреді. Сонымен тыныштық потенциалы калий иондарының циоплазмадан сыртқы ортаға қарай бағытталған диффузия әсерінен деп қарастырсақ, онда оның шамасын Нернст
теңдеуінен анықтауға болады:
j= RT/nF ln C1/C2
мұндағы К1 жасушаның ішіндегі, К2 сыртындағы калии ионының концентрациясы. Бұл теңдеуді тепе теңдік потенциалы деп те атайды. Кальмар аксонындағы тыныштық потенциалының шамасы jМ = - 60мВ (2 сурет).
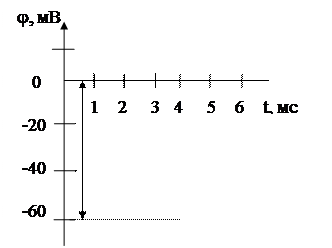 |
jМ = - 60мВ
2 сурет
Кейінгі жүргізілген тәжірибелер мембрананың аз мөлшерде натрия мен хлор иондарын да өткізетіндігін көрсетті, олай болса тыныштық потенциалының пайда болуында K+, Na+ және Cl- иондарынының да қатысы бар. Қозбаған мембрананың K+, Na+ және Cl- иондары өткізу коэффиценті PK : PNa : PCl = 1 : 0,04 : 0,45 мұнан калий ионының натриға салыстырғанда өтімділігі 25 есе жоғары екендігі көрінеді, яғни тыныштық потенциалының пайда болуында калии мен натрии иондарының концентрациялары басты роль атқаратындығы келіп шығады.
1949 жылы Гольдман, Ходжкин және Катц тыныштық потенциалының басты теңдеуін ұсынды:
jM = -RT/F ln 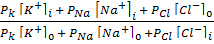 ,
,
мұндағы i и 0 иондардың ішкі және сыртқы ортадағы концентрациясы, РК, РNa РCl сәйкес иондар үшін мембрананың өткізгіштік коэффиценті. Бұл өрнек Гольдман- Ходжкин - Катц теңдеуі деп аталады.
Жасушаны сыртқы тітіркендіргіштер (механикалық, жылулық, электрлік) арқылы қозған күйге көшірсек, мембрананың натрии иондарын өткізуі күрт жоғарылайды, бірақ калии иондарын өткізу дәрежесі өзгеріссіз қалады. Мембрананың калии және натрии иондарын өткізу коэффициенті PK : PNa = 1 : 20 дейін өзгеріп, натрии ионын өткізу 500 есе артады. Оның үстіне мембранада натрии иондарының каналы 10 есе көп, осы факторлар натрии иондарының мембрана ішіне қарай өтуін күшейтіп, ондағы потенциалдар айрымын төмендеуіне, соңынан оның ішкі қабатының оң зарядталуына алып келеді. Жасушадағы бұл құбылысты деполяризация деп атайды. Мембрананың қозу нәтижесіндегі потенциалдар айрымының толық өзгерісі әрекет потенциалының мәнін береді: jӘ = jТ - Dj, где jТ – тыныштық потенциаланың мәні, Dj- потенциал өзгірісі. Әрекет потенциалы деп қозған күйдегі цитоплазма мен сыртқы орта арасындағы потенциалдар айрымын айтамыз. Мысалы, үлкен кальмар аксонының тыныштық потенциалы -60 Мв тең. Оны қоздырғанда потенцилы + 35 мВ дейін жоғарыласа, онда потенцилдың айырымы немесе әрекет потенциалы jӘ = - 60 мВ – 35 мВ = - 95 мВ ( 3- сурет).
Дата добавления: 2020-11-18; просмотров: 686;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
