Основные сведения о строении и физиологии нервной системы человека
Клинические аспекты логопедии являются основными при изучении и оказании педагогической помощи лицам с синдромами нарушений речи, возникающих при поражении того или иного участка мозга. В этих целях необходима качественная оценка признаков речевой патологии в сопоставлении с локализацией повреждения ЦНС. Такой анализ позволяет психолого-педагогическим методам реабилитации быть более оправданными и целенаправленными.
С развитием нейробиологии - науки о строении и функции нервной системы человека - во многом становятся понятными особенности развития организма человека и его речи.
В основе современного представления о структуре и функции центральной нервной системы (ЦНС) лежит нейронная теория, согласно которой мозг представляет собой функциональное объединение отдельных клеточных элементов - нейронов.
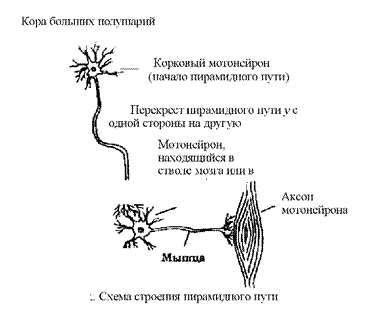
Через пирамидный (кортиконуклеарный) путь осуществляется регуляция произвольных движений моторными отделами коры, а значит, непосредственно обеспечивает произвольные движения.
Нейрон - это нервная клетка, обладающая способностью генерировать возбуждение в ответ на раздражение и передавать его другим нейронам или эффекторным органам (мышцам, железам). По функции нейроны разделяются на: афферентные (чувствительные), эфферентные (двигательные), вставочные.
Аксоны афферентных нейронов воспринимают сигналы, возникающие в рецепторных окончаниях органов чувств (мышц в т.ч.), и проводят их в ЦНС.
Аксоны эфферентных нейронов, например, выходят за пределы ЦНС и иннервируют скелетную мускулатуру. Многие эфферентные нейроны передают сигналы к органам и мышцам посредством других нервных клеток и их отростков (например, от пирамидных клеток моторной области коры импульсы поступают к мотонейронам продолговатого и спинного мозга и далее по периферическим нервам к мышцам).
Вставочные нейроны обеспечивают связь между афферентными и эфферентными нейронами.
Нейроны. в т. ч. моторные ядра, или мотонейроны, являясь клеточными элементами ЦНС, обладают особым видом спонтанной электрической активности, имеющей эндогенное происхождение. Эта врожденная ритмическая активность превращает нейрон в генератор возбуждения исмекерный механизм). Уровень эндогенного возбуждения нейрона может повышаться и снижаться в зависимости от афферентных воздействий на нейрон. Таким образом, пейсмекерная активность мотонейронов черепно-мозговых нервов и периферических нервов спинного мозга существенно зависит от влияния со стороны корковых нейронов. Чем меньше сила влияния корковых отделов мозга, тем выше пейсмекерная активность нейронов, находящихся в продолговатом и спинном мозге.
В зрелом состоянии головной и спинной мозг, а также весь аппарат периферических нервов с рецепторными органами (т.е. органами, в которых заканчиваются нервные окончания) представляют целостную систему, которая анатомически и функционально делится на большое число звеньев. ЦНС включает в себя те части нервной системы, которые лежат внутри черепа и позвоночного столба. Нервы, лежащие вне черепа или позвоночника, представляют периферическую нервную систему. ЦНС подразделяется на спинной и головной мозг (рис. 2).
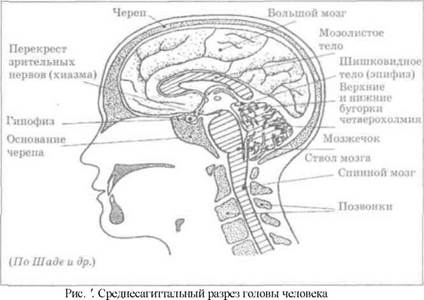
Спинной мозг является цилиндрическим образованием, которое состоит из серого вещества тел нейронов, сконцентрированных в центральной части мозга в виде «бабочки». Передние рога «бабочки» содержат нейроны, эфферентные аксоны которых направляются в составе спинно-мозговых нервов к мышцам, т.е. в составе периферических нервов. Задние рога содержат клетки промежуточных нейронов, к которым подходят афферентные волокна, входящие в состав периферических нервов и доставляющих чувствительные импульсы с периферии. Спинно-мозговые нервы уровня шейного отдела позвоночника иннервируют диафрагму.
 |  | ||
Головной мозг подразделяется на задний, средний, промежуточный и передний мозг.
Задний мозг состоит из продолговатого мозга, мозжечка и варолиева моста.
Продолговатый мозг является нижним отделом головного мозга.
На передней поверхности продолговатого мозга имеются два вертикальных валика, получивших название пирамид. По бокам от них расположена другая пара валиков, обозначаемых как оливы.
Продолговатый мозг включает ряд ядер, которые обслуживают сосудисто-двигательный и дыхательный центр, контролирующие сужение и расширение сосудов, а также сердечный ритм. Ядра продолговатого мозга принимают участие в обеспечении таких сложных рефлекторных актов, как сосание, жевание, глотание, рвоты, чихание, моргание (функции блуждающего, языкоглоточного, подъязычного и тройничного нервов).
Центры продолговатого мозга в процессе эмбриогенеза развиваются и созревают раньше, чем другие отделы мозга. О сохранности центров продолговатого мозга в процессе внутриутробного развития свидетельствует крик ребенка непосредственно в процессе родов. Наличие регуляторных функций дыхания и сердца, сосательного рефлекса в первые минуты и часы жизни. Нарушение этих функций у ребенка при рождении свидетельствует о поражении ствола мозга.
В мозжечке различают два полушария и его центральную часть - червь.
Мозжечок обеспечивает точность целенаправленных движений, координирует действия мышц - антогонистов (мышц противоположного действия), регулирует мышечный тонус, поддерживает равновесие.
Обеспечение этих функций осуществляется благодаря тесным связям мозжечка со всеми структурами мозга: с чувствительной сферой (проприорецепция - положение туловища в пространстве), с экстрапирамидной системой, с ретикулярной формацией ствола, с лобными, затылочными и височными отделами коры головного мозга.
Мозжечок имеет тесные связи. Корково-мостомозжечковые пути (лобно-мостомозжечковый и затылочно-мостомозжечковый) проходят из коры головного мозга к собственным ядрам Варолиева моста, а затем к коре мозжечка противоположной стороны. Эти импульсы корригируют деятельность экстрапирамидной системы мозжечка - это нисходящие пути. Восходящие пути, которые несут проприоцептивную информацию в мозжечок, сформированы в переднеспинно-мозжечковый путь (начинается в проприорецепторах и достигает червя мозжечка) и заднеспинно-мозжечковый (путь также начинается в проприорецепторах и также достигает червя мозжечка).
Таким образом, мозжечок вместе с лобной корой больших полушарий соучаствует в формировании моторных программ на основе импульсов, поступающих в него от мышечно-суставных проприорецепторов, а также от вестибулярных зрительных и тактильных анализаторов.
Варолиев мост представляет собой крупное поперечно-волокнистое образование, охватывающее передневерхнюю часть продолговатого мозга. Здесь находятся ядра: двигательное ядро отводящего нерва (VI пара), двигательное ядро тройничного нерва (V пара), два чувствительных ядра тройничного нерва ядра слухового и вестибулярного нервов, ядро лицевого нерва (VII пара), собственные ядра моста, в которых переключаются корковые пути, идущие в мозжечок.
Через Варолиев мост проходит большое количество проводящих путей. К основным относятся: двигательный кортиконуклеарный (пирамидный) от коры к мозжечку и общий чувствительный путь.
В функцию моста входит проведение координирующих импульсов от одного полушария мозжечка к другому для обеспечения согласованных сокращений/расслаблений мышц с обеих сторон тела и связи коры больших полушарий с самим мозжечком.
Средний мозг расположен под мостом и включает ножки мозга и четверохолмие. В каждой из ножек мозга, помимо проводящих волокон, располагаются богатые пигментом черная субстанция и красное ядро, имеющие отношение к регуляции движений. Черное вещество и красное ядро являются частью паллидарной системы. Вместе с ретикулярной формацией принимает участие в регуляции мышечного тонуса при выполнении точных и плавных движений пальцев рук.
Важнейшей функцией этого отдела мозга является перераспределение мышечного тонуса. Он участвует в реализации статических рефлексов (положение тела в пространстве) и статокинетических рефлексов (перемещение тела).
Промежуточный мозг включает два отдела: зрительный бугор (таламус) и подбугорную область (гипоталамус).
Таламус выполняет функцию коллектора и коммутатора всех возбуждений, поступающих от рецепторов в головной мозг (кроме обоняния), т.е. производит предварительный анализ и синтез импульсов от всех органов чувств и через синаптические связи направляет их в различные зоны мозга, в том числе коры больших полушарий.
Гипоталамус представляет собой относительно узкий слой мозговой ткани. В нем расположены многочисленные высокодифференцированные ядра, регулирующие температуру тела, аппетит, водный баланс, углеводный и жировой обмены, сосудистый тонус и другие вегетативные функции, которые связаны с обменом веществ. Здесь же находятся центры, осуществляющие регуляцию сна, сексуального и эмоционального поведения. Гипоталамус играет важнейшую роль в регуляции постоянства внутренней среды организма (гомеостаза). Кроме того, к структурам гипоталамуса анатомически относят гипофиз - железу внутренней секреции и зрительную хиазму - место неполного перекрестья зрительных нервов.
Все отделы между спинным мозгом и промежуточным мозгом образуют ствол мозга, т.е. в его состав входят средний мозг, варолиев мост, продолговатый мозг. Мозговой ствол является промежуточной инстанцией. В нем проходят из спинного мозга в большой мозг афферентные волокна, а из большого мозга к передним рогам спинного мозга эфферентные волокна. В стволе находятся ядра III и XII пар черепно-мозговых нервов, а также ядерные образования экстрапирамидной системы. Здесь имеются важнейшие центры вегетативной иннервации, которые контролируют дыхание и сердечно-сосудистую деятельность.
В массе ствола мозга расположена ретикулярная (сетевидная) формация, волокна которой переплетаются со всеми проходящими через ствол мозга афферентными и двигательными путями. Ядерные образования ретикулярной формации, их многочисленные нейроны дают начало эфферентным связям, которые подразделяются на: нисходящие и восходящие.
Нисходящая система состоит из активирующих и тормозных волокон, которые регулируют деятельность спинного мозга. Активирующее влияние проявляется повышением мышечного тонуса, а тормозящее - снижением тонуса мышц.
Анатомо-фтиологические и психофизиологические основы речевой функции
Основная масса клеток ретикулярной формации образует систему вставочных нейронов, которые обеспечивают совместную координированную деятельность различных отделов нервной системы.
Она оказывает энергорегулирующее воздействие на кору больших полушарий и контролирует рефлекторную деятельность мозга.
Передний мозг состоит из двух полушарий, покрытых серым веществом - корой. У человека под влиянием социальной среды в процессе онтогенеза формируются особые структуры коры больших полушарий мозга, наиболее дифференцированный отдел центральной нервной системы. В самой нижней части премоторной извилины левого полушария (главным образом, у правшей) расположена височная область, в центре которой находится зона Брока, которая реализует двигательную сторону речи. На заднем участке височной извилины, на стыке первичной слуховой и двигательной коры находится зона Вернике, реализующая восприятие речи.
Многочисленные современные данные свидетельствуют о том, что организация речи осуществляется при взаимодополняющем постоянном взаимодействии двух полушарий.
Нейроанатомические различия правого и левого полушария отмечены как в речевых зонах, так и в других структурах, в первую очередь затылочных и верхнетеменных. Три основные модальности (ощущение звука, света, осязание) наиболее представлены в левой гемисфере (у правшей). С разными полушариями связан и характер эмоционального реагирования. Асимметрия эмоциональной сферы выражается в преимущественной «ответственности» левого полушария за формирование положительных эмоций, а правого - отрицательных. Повреждение левого полушария на ранних этапах онтогенеза не приводит к речевым расстройствам (например, алалии), так как в правом полушарии имеются нейроанатомические предпосылки для развития «речевых» зон. В то же время при поражении правого полушария нарушаются невербальные психические функции, которые не компенсируются левым полушарием.
С деятельностью правого полушария связывают регулирование активности речевых центров левого полушария, обеспечивание помехоустойчивости речевого слуха, интонационные характеристики речи, конкретность и предметность высказываний. С деятельностью левого полушария связаны главным образом языковые уровни: фонологическая система языка, морфологический механизм словообразования, синтаксическое структурирование высказывания, кратковременная и долговременная словесная память. Отсюда понятно положительное воздействие на динамику речевого развития при адекватном сенсорном воспитании.
В процессе реализации любой задачи, требующей сенсорного или моторного решения (например, чтение, письмо или любое другое произвольное действие), вовлекаются в деятельность структуры обоих полушарий. Следует более четко представлять то, что межполушарная асимметрия в деятельности мозга имеет сложный характер, который до настоящего времени полностью не изучен.
В глубине каждого полушария расположены проводящие волокна и подкорковые ядра (базальные ганглии). Наиболее крупным образованием является полосатое тело (стриатум), которое состоит из хвостатого ядра, скорлупы и бледного шара (паллидум). Эти ядра объединяются общим названием - стриопаллидарная система. За счет стриопаллидарной системы у новорожденного осуществляются диффузные массовые движения тела (рис. 3).
После созревания моторных областей коры больших полушарий стриопаллидарная система начинает обеспечивать «готовность» к совершенствованию движений, а именно, перераспределяет и согласует тонус мышц, что позволяет произвольным движениям быть быстрыми, точными и строго дифференцированными.
Со стриопаллидарной системой, являющейся эфферентным звеном в нервной системе, функционально тесно связан зрительный бугор (таламус). На уровне таламуса происходит формирование сложных рефлексов смеха и плача.
Каждое полушарие головного мозга разделено глубокими бороздами на большие участки, называемые долями. Такими бороздами являются: боковая (сильвиева борозда), центральная (роландова борозда) и теменно-затылочная борозда. Продольная щель мозга делит его на два полушария. Каждое полушарие состоит из пяти долей: лобная, теменная, височная, затылочная и долька, скрытая на дне сильвиевой борозды - островок.
Оба полушария объединены между собой спайками, наиболее крупная из которых мозолистое тело, которое расположено выше таламуса.
Совокупность отделов мозга, включающая внутреннюю поверхность полушарий (медиобазальные отделы) и их глубокие структуры, получила название лимбической системы. Традиционно в лимбическую систему включают миндалевидное тело, гипоталамус и ретикулярную формацию среднего мозга, объединенные под названием - глубинные структуры мозга. Особенностью лимбической системы является то, что между составляющими ее структурами имеются простые двусторонние связи и сложные пути, образующие множества замкнутых кругов. Такая организация создает условия для длительного циркулирования одного и того же возбуждения в системе и, тем самым, сохранение в ней единого состояния, а также навязывания его другим системам мозга. Круги возбуждений разного функционального назначения связывают лимбическую систему со многими структурами большого мозга. Подавляющее большинство структур лимбической системы принимает участие в функциональной организации эмоций, что предполагает их влияние на соответствующие вегетативные изменения, регулируемые гипоталамусом.
Кора головного мозга является наиболее дифференцированным отделом ЦНС, состоящим из шести слоев нейронов разного типа. Для коры характерно обилие межнейрон пых связей. Особо многочисленные контакты имеются в сенсо-моторных отделах, что позволяет координировать моторную функцию с разнообразными воздействиями как из внешней, так и внутренней среды организма.
Различные структуры мозга созревают неравномерно, и поэтому готовность к выполнению свойственных им функций наступает в разные возрастные периоды. Сначала происходит созревание глубоких структур, подкорковых образований, отвечающих за принципиальные стороны жизнедеятельности. Затем созревают так называемые первичные зоны мозга, в которых заканчиваются нервные волокна, идущие от периферических частей анализаторов. Те и другие являются почти сформированными к моменту рождения. В первый год жизни они функционально оформляются, создавая основу сенсомоторной стадии развития.
Созревание ассоциативных зон мозга осуществляется в период от двух до пяти лет. В последнюю очередь развиваются лобные доли мозга, морфологическое структурирование которых происходит в возрасте от 12 до 14 лет.
Моторика представляет собой всю сферу двигательных функций организма, включающая биомеханические, физиологические и психологические аспекты.
Движения являются главным средством взаимодействия организма человека с окружающей средой. Основные типы движений человека сводятся к четырем типам активности: обеспечение позы и равновесия, локомоция и произвольные движения. Произвольными движениями могут быть названы разнообразные двигательные акты, совершаемые человеком в процессе повседневном жизни. Они являются целесообразными и носят навыковый (условно-рефлекторный) характер, в отличие от врожденных, безусловно-рефлекторных моторных реакций (чихание, глотание и т.д.). ЦНС снабжается информацией о состоянии периферического двигательного аппарата с помощью разного вида чувствительных нервных окончаний (рецепторов), которые обеспечивают т.н. проприо-цептивную чувствительность.
К центральному механизму произвольных движений относится кортиконуклеарныи (пирамидный) путь, который идет от двигательной зоны коры головного мозга и заканчивается в ядрах ствола к двигательным ядрам черепно-мозговых нервов и ядрах спинного мозга (мотонейроны спинного мозга).
Праксис - это такой функциональный уровень в организац произвольных действий, где их координация обусловливается (афферентируется) обобщенными топологическими признаками объекта, т.е. смысловой последовательностью элементов движения (застегнуть пальто, налить воды в чашку и т.п.). Можно назвать это символичным уровнем движений (Н.А. Бернштейн, 1946).
В корковом отделе речедвигательного анализатора доминантного полушария имеются зоны, обеспечивающие подготовку отдельных сторон речедвигательного акта:
– оценка исходного положения органа периферического аппарата (по сумме кинестетических импульсаций);
– организация речедвигательного акта во времени и как планирования серии последовательных сокращений отдельных мышечных групп этого органа (например, отдельных групп мышц языка);
– программа речедвигательного акта в пространстве движения отдельного органа (например, подъем кончика языка к альвеолам).
Эти три программы осуществляются в трех различных зовах коркового речедвигательного анализатора.
По А.Р. Лурия, в реализацию двигательного акта, помимо собственно моторных зон, включается почти вся кора. Передние отделы мозга связаны с построением кинетических программ двигательного акта, а задние - с их кинестетическим и пространственно-обусловленным обеспечением.
Кинестетический фактор обеспечивает передачу и интеграцию сигналов, поступающих от рецепторов, расположенных в мышцах, суставах и сухожилиях, которые несут информацию о взаимном расположении органов. Эти сведения принимает передняя часть теменной области, куда притекают также тактильные и зрительные импульсы, что делает информацию полной. Исключение составляет речевая артикуляция, которая функционирует на кинестетической основе без участия зрения (тем не менее, в онтогенезе дети с нормальным зрением начинают говорить раньше, чем дети с нарушенным зрением). Информацию о речевой моторике, помимо кинестезии, у лиц с развитой речью дает акустический контроль.
Кинетический фактор реализуется в двигательных актах, которые осуществляются в форме кинетических и мелодических схем. В отношении речевой моторики этот фактор обеспечивает плавную смену артикуляции в процессе произнесения и перехода от звука к звуку в слове, от слова к слову. При нарушении кинетического фактора возникают «застревания» на фрагментах движения, что приводит к неоднократному повторению этого фрагмента. В устной речи это проявляется в повторении звуков и слогов, букв и их элементов при письме.
Конкретные эфферентные механизмы исполнения движений обеспечиваются пирамидной и экстрапирамидной системами, корковые отделы которых составляют единую сенсо-моторную зону коры (рис. 4).
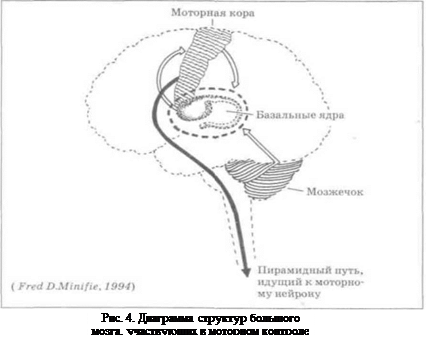
Пирамидная система (центральный двигательный путь) участвует в организации точных пространственно-ориентированных движений и полностью подчинена произвольному контролю. Клетки центральных двигательных невронов сосредоточены главным образом в передних центральных извилинах, а также в теменных долях коры. Кортиконуклеарный путь заканчивается в моторных ядрах черепно-мозговых нервов, которые находятся в варолиевом мосту, продолговатом спинном мозге. На границе продолговатого и спинного мозга большая часть волокон пирамидного пути правого и левого полушария перекрещивается. Ядра тройничного, языкоглоточного, блуждающего и часть ядра лицевого нерва получают импульсы от обоих полушарий мозга, так как подходящие к ним волокна пирамидного пути перекрещиваются неполностью. Именно этим объясняется то, что односторонние очаговые поражения пирамидных путей не вызывают серьезных функциональных нарушений жевания, глотания и голосообразования. Исключение составляют волокна, несущие корковые импульсы к ядру подъязычного нерва, которые полностью перекрещиваются в продолговатом мозгу, непосредственно перед вступлением их в ядро. Поэтому ь чае поражения мозга на уровне варолиева моста и выше спастический паралич мышц языка наблюдается на стороне, противоположной очагу поражения (т.е. при этом язык отклоняется в сторону очага поражения). Остальная часть волокон заканчивается в моторных ядрах спинного мозга.
Экстрапирамидная система обеспечивает автоматизированные движения. В основном она управляет непроизвольным компонентом движений: поддержание позы, физиологические синергии, общую согласованность двигательных актов, их пластичность. Традиционно в ней различают корковый и подкорковый отделы (стриопаллидарная система, красное ядро и черная субстанция, мозжечок и ретикулярная формация ствола мозга и их корковые отделы).
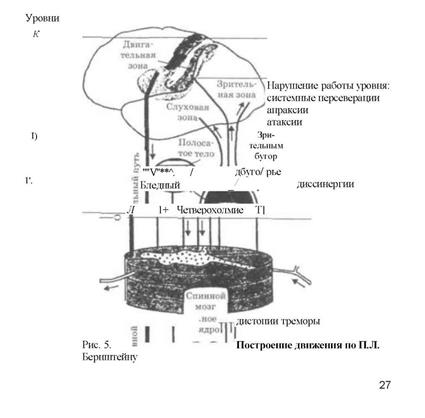
Результаты исследования движений позволили Н.А. Бернштейну (1965) сформулировать общие представления и многоуровневой иерархической системе координации движении. В соответствии с ними система управления движениями состоит из следующих уровней: А - уровень палеокинетических регуляций, он же руброспинальный уровень ЦНС. Действия этого уровня полностью непроизвольны; В - уровень синергии, он же таламопаллидарный уровень. Движения этого уровня характеризуются стереотипностью, в обобщенном виде это афферентация собственного тела. Этот уровень обеспечивает такие врожденные особенности моторики, как ловкость, грациозность, пластику. Нарушения этого уровня ведут к насильственным движениям; С - уровень пространственного поля, он же пирамидно-стриарный. Этот уровень обеспечивает все переместительные движения: ходьба, прыжки и т.д. Патология этого уровня сопровождается нарушениями пространственной координации (атоксия), равновесия, локомоции и точности; D - теменно-премоторный или уровень предметных действий, которые не являются врожденными, а формируются и совершенствуются в процессе накопления опыта; Е - группа высших кортикальных уровней символических координации (письма, речи и т.д.).
На рис. 5 представлена схема основных центров и проводящих путей мозга с распределением их по уровням.
А.Р. Лурия (1969) разработал общую структурно-функциональную модель мозга как органа, с которым связана психомоторная деятельность человека. Работа мозга обеспечивается тремя блоками, которые характеризуются особенностями строения и участия в реализации психических функций.
I блок - энергетический. Он включает ретикулярную формацию, ствол мозга, неспецифические структуры среднего мозга, лимбическую систему, медиобазальные отделы коры лобных и височных долей. Блок регулирует «тонус мозга», необходимый для выполнения психической деятельности, т.е. он поддерживает бодрствующее состояние и сознание в целом.
II блок - блок приема, переработки и хранения экстеро-цептнвной (внешней) информации. В него включаются центральные части основных анализаторных систем: зрительной, слуховой и кожно-кинестетической. Корковые зоны этих анализаторных систем расположены в затылочных, теменных и височных долях мозга (первичные корковые поля). Эти зоны коры называются проекционными.
Вторичные корковые поля представляют клеточные структуры, в которых происходит усложнение переработки первичной информации благодаря прохождению афферентных импульсов через ассоциативные ядра таламуса. Их функциональная организация на уровне психики эквивалентна процессу восприятия.
Первичные и вторичные поля относятся к ядерным зонам анализаторов, их нейроны модально специализированы.
Третичные поля - ассоциативные; расположены на границе затылочных, височных и заднецентральных отделов коры. Их функция состоит в интеграции возбуждений, приходящих от вторичных полей всего комплекса анализаторов. Это обеспечивает возможность реакции третичных полей на обобщенные признаки объектов и явлений.
III блок - блок программирования, регуляции и контроля за протеканием психической сознательной деятельности. Он включает моторные, премоторные и нрефронтальные отделы коры лобных долей мозга. Основная функция этого блока представляет собой программирование психического акта и развертки последовательности его реализации во времени.
Анатомо-физиологические и психофизиологические основы речевой функции Зоны иннервации черепно-мозговых нервов
| 3, 4, 6 пары | Глазодвигательные вервы | Мышцы глазного яблока |
| 5 пара | Тройничный нерв | 1 ветвь - кожа лба, верхнего века, слизистая оболочка полости носа; 2 ветвь - кожа нижнего века, носа, верхней губы; слизистая щек, верней губы; верхние зубы, десна; 3 ветвь - кожа нижней части лица, слюнные железы, нижние зубы и десна: мышцы передней части языка, жевательные мыш цы |
| 7 пара | Лицевой нерв | Мимические мышцы лица, кожа головы, мышцы затылочной области, ушной раковины, частично дна полости рта; подкожная мышца шеи |
| 8 пара | Слуховестибуляриый нерв | Органы слуха |
| 9 пара | Языкоглоточный нерв | Слизистая оболочка глотки, надгортанника; мышцы корня языка, мягкого неба |
| 10 пара | Блуждающий нерв | Мышцы передней части языка, надгортанника |
| 1 1 пара | Добавочный нерв | Грудиноключично-сосцевидную и трапециевидную мышцы |
| 12 пара | Подъязычный нерв | Мышцы языка |
Работа каждого блока не является автономной, а представляет собой результат координированного взаимодействия всех трех структур. Итогом деятельности этой системы является нерасчленяемая и неосознаваемая моторная активность и в целом психическая жизнь. Эти положения А.Р. Лурия полностью относятся к так называемым речевым структурам. Было установлено, что при возникновении патологии в различных участках коры, связанных с речью, их функции берут на себя сохранившиеся отделы как левого, так и правого полушария. Таким образом, речевые структуры мозга обладают широкой распределенностью и полифункциональностью, определяемой возможностью их полного взаимодействия. Однако существует обязательное звено, без которого осуществление речевого акта становится невозможным. Среди прочих речевых структур у большинства взрослых людей таким звеном является левополушарная кора, при нарушении которой возникает афазия и корковая дизартрия (по Е.Н. Винарской, 2005).
Дата добавления: 2016-07-27; просмотров: 3552;
Поиск по сайту
Узнать еще
- Arthropoda.Систематика.Тараканы и мухи.Географическое распространение.Основные представители.Морфология,развитие,патогенное действие.Медицинское знаение.Меры борьбы.
- Cимпатическая нервная система. Центральный и периферический отдел симпатической нервной системы.
- Cущность организации и ее основные признаки
- I. Назначение унифицированных газозарядных станций и основные тактико-технические требования, предъявляемые к ним.
- I. ОСНОВНЫЕ ПОЛОЖЕНИЯ
- I. Политический режим: понятие, сущность и основные типы.
- I.2. Антигены системы АВ0. Генетика. Структура
- I.2. Основные категории водопотребления промышленных предприятий и их особенности

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
