Ферменты-рестриктазы и рестрикция ДНК
В ходе эволюции бактерии развили способность синтезировать так называемые рестрицирующие ферменты (эндонуклеазы), которые стали частью клеточной (бактериальной) системы рестрикции-модификации. У бактерий системы рестрикции-модификации являются внутриклеточной иммунной системой защиты от чужеродной ДНК. В отличие от высших организмов, у которых распознание и разрушение вирусов, бактерий и других патогенов происходит внеклеточно, у бактерий защита от чужеродной ДНК (ДНК растений и животных, в организме которых они обитают) происходит внутриклеточно, т. е. тогда, когда чужеродная ДНК проникает в цитоплазму бактерий. С целью защиты бактерии в ходе эволюции развили также способность «метить» собственную ДНК метилирующими основаниями на определенных последовательностях. По этой причине чужеродная ДНК из-за отсутствия в ней метальных групп на тех же последовательностях плавится (разрезается) на фрагменты разными бактериальными рестриктазами, а затем деградируется бактериальными экзонуклеазами до нуклео-тидов. Можно сказать, что таким образом бактерии защищают себя от ДНК растений и животных, в организме которых они обитают временно (как патогены) или постоянно (как сапрофиты).
Рестриктазы впервые были выделены из Е. coli в 1968 г- Оказалось, что они способны разрезать (плавить) молекулы ДНК на разных сайтах (местах) рестрикции. Эти ферменты получили название эндонуклеаз класса I. Затем у бактерий были обнаружены эндонуклеазы класса II, которые распознают в чужеродной ДНК сайты рестрикции специфически и на этих сайтах тоже осуществляют рестрикцию. Именно ферменты этого класса стали использовать в генной инженерии. Тогда же были открыты ферменты класса III, которые плавят ДНК рядом с сайтами распознания, но эти ферменты не имеют значения в генной инженерии.
Действие системы рестрикции-модификации «рационализуется» так называемыми палиндромными (распознающими последовательностями) азотистых оснований, которые являются сайтами рестрикции ДНК. Палиндромные последовательности — это последовательности оснований, которые одинаково читаются вперед и назад, как, например, последовательность букв радар. Поскольку цепи ДНК обладают антипараллельным направлением, то считают, что последовательность является палиндромной, если она идентична, когда читается в направлении от 5'- к 3'-концу на верхней и 3'- к о'-концу на нижней цепи, а именно:
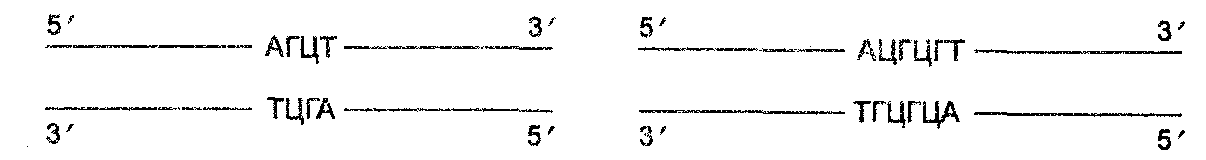
Палиндромы могут быть любых размеров, но большинство тех палиндромов, которые используют в качестве сайтов узнавания рестриктазами, состоят из 4, 5, 6 и реже 8 оснований.
Рестриктазы — это абсолютно необходимый инструмент в генной инженерии для вырезания интересующих фрагментов (генов) из больших молекул ДНК. Поскольку известно более 100 ферментов рестрикции, то это позволяет выбор рестриктаз и селективное вырезание фрагментов из исходной ДНК.
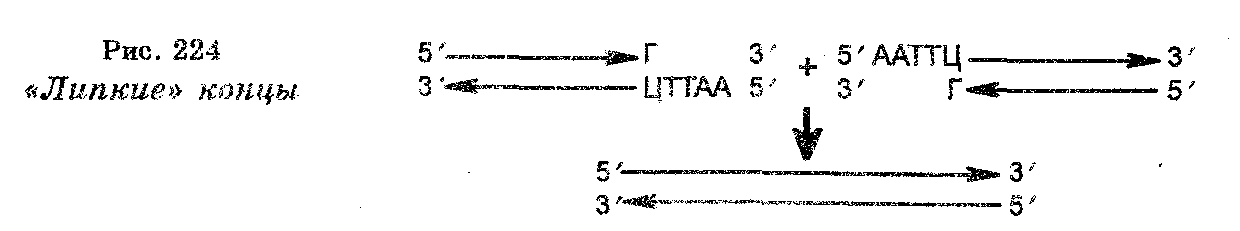
Замечательной особенностью рестриктаа является то, что они продуцируют разрезы молекул на несколько фрагментов (рестрик-тов) ДНК уступами, в результате чего в образующихся концах одна цепь длиннее другой, образуя своеобразный хвост. Такие концы (хвосты) получили название «липких» концов (рис. 224), т. к. они способны к самокомплементарности.
Рассмотрим результаты рестрикции на примере одной из наиболее известных рестриктаз Eco RI из системы рестрикция—модификация Е. coli. Вместо того, чтобы плавить ДНК в центре па-линдромной последовательности узнавания, этот фермент плавит ДНК за пределами центра и продуцирует 4 самокомплементарных («липких») конца, состоящих из разного количества нуклео-тидов, а именно:
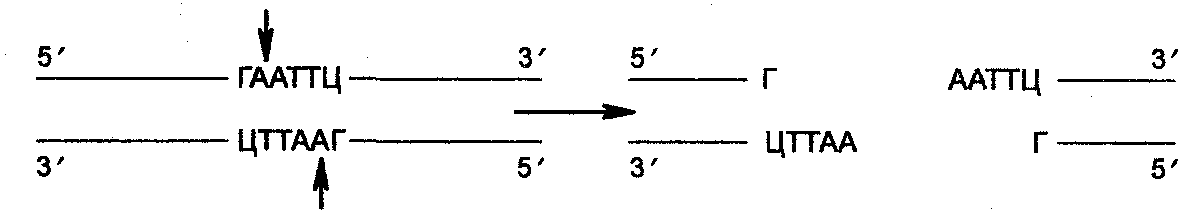
Эти «липкие» концы в генно-инженерных опытах полезны по той причине, что они могут быть воссоединены комплементарно при низких температурах, что позволяет эффективное смыкание ДНК-фрагментов.
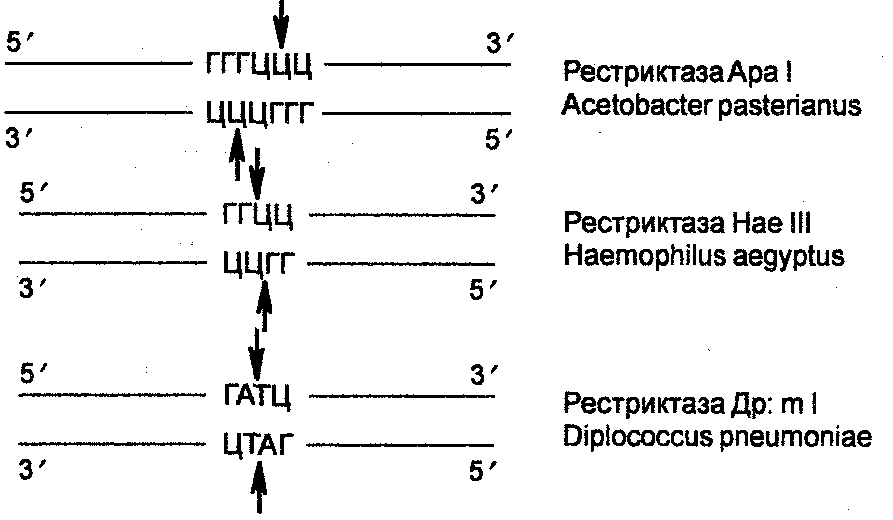
Сайты распознания и сайты плавления в случае других рестриктаз имеют другое содержание, а именно:
Вслед за рестрикцией ДНК из рестрикционной смеси выделяют рестрикционные ДНК-фрагменты (ДНК-рестрикты), которые необходимы затем для объединения с вектором. Для выделения ДНК-рестриктов прибегают к электрофорезу, поскольку с помощью этого метода рестрикцированную ДНК очень легко фракционировать благодаря размерам фрагментов-рестриктов и благодаря константным отношениям электрический заряд-масса. Фрагменты в электрическом поле мигрируют в ходе электрофореза при частоте, зависимой от их размеров (массы). Чем больше (длиннее) фрагмент, тем медленнее он мигрирует в электрическом поле. Материалом, в котором проводят электрофорез, являются незаряжающиеся агаро-за и полиакриламид. Для опознания фрагментов используют эти-дий бромид, который красит фрагменты, что ведет к их более легкому обнаружению (рис. 225).
 Результативность электрофореза очень высока, поскольку с его помощью могут быть разделены фрагменты, размеры которых составляют от 2 до 50 000 оснований.
Результативность электрофореза очень высока, поскольку с его помощью могут быть разделены фрагменты, размеры которых составляют от 2 до 50 000 оснований.
После электрофореза фрагменты из агарозы выделяют с помощью разных методов. На основании результатов сравнения размеров рестриктов одной и той же ДНК, полученных с помощью разных рестриктаз, строят рестрикционные карты, на которых показывают сайты рестрикции каждой из использованных рестриктаз (см. § 107). В практическом плане рестрикционные карты позволяют определять не только размеры рестриктов, но и выяснять расположение в молекулах ДНК локусов тех или иных генов.
Поскольку у высших организмов в ходе транскрипции синтезируется гетерогенная ДНК, корректируемая процессингом, то в генной инженерии обычно используют комплементарную ДНК (кДНК), которую получают при использовании в качестве матрицы мРНК, на которой обратная транскриптаза синтезирует одноцепочечную ДНК (кДНК), являющуюся копией мРНК. В последующем эти одноцепочечные ДНК превращают в двухцепочечные ДНК. Считают, что кДНК содержит непрерывные нуклеотидные последовательности (транскрибируемые и транслируемые). Именно кДНК используют для рестрикции.
Выделенные после электрофореза из агарозных гелей фрагменты ДНК (рестрикты) можно предварительно подвергнуть секвени-рованию, т. е. определить в них нуклеотидную последовательность. Для этого используют химический и ферментативный методы сек-венирования.
Химический метод основан на получении меченных радиоактивным фосфором (32р) фрагментов и удалении из этих фрагментов одного из оснований с последующим учетом результатов радиоавтографии гелей, содержащих эти фрагменты. Ферментативный метод основан на том, что в конец анализируемого фрагмента вводят нуклеотид, используемый затем в синтезе разных фрагментов in vitro, анализируемых на нуклеотидную последовательность электрофоретически. Для изучения специфических последовательностей нуклеотидов в молекуле ДНК используют также гибридизацию ДНК-ДНК, РНК-РНК, ДНК-РНК, Нозерн- и Саузерн-блоттинги.
Дата добавления: 2016-05-30; просмотров: 2713;
Поиск по сайту
Узнать еще
- Альтернативные структуры ДНК
- Биосинтез белков, код ДНК, транскрипция
- В короткой интерфазе не происходит репликации ДНК. К полюсам клетки расходятся хроматиды
- Введение рекомбинантной ДНК в клетки бактерий и отбор трансформантов
- Векторы для клонирования ДНК
- Векторы, используемые для клонирования ДНК
- Встраивание ДНК в вектор
- Вторичная структура ДНК. Правила Чаргаффа.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
