Ограничение и усвоение фосфора донными водорослями: ключевые процессы и методы оценки
Повышенное поступление биогенных элементов в ручьи и озёра представляет серьёзную экологическую угрозу. Загрязнение из диффузных источников, обусловленное изменением землепользования, приводит к ухудшению состояния водных объектов (Джордан и др., 1997; Карпентер и др., 1998; Аллан, 2004). Среди основных нарушений выделяют культурную эвтрофикацию, вредное цветение водорослей, тепловое загрязнение, аккумуляцию осадков и накопление пестицидов, нефтепродуктов. Устойчивость водной экосистемы к этим стрессорам определяется её биологическими, химическими, геологическими и геоморфологическими характеристиками. В речных экосистемах донные водоросли и бактерии выступают важным биотическим поглотителем загрязняющих веществ, включая избыток биогенов (Малхолланд и Роземонд, 1992; Бернхардт и др., 2003).
Определение скоростей поглощения и выделения фосфора (Р) даёт информацию о предельно допустимой нагрузке на ручей, озеро или эстуарий до потери их экологической целостности (Доддс, 2003). Именно неорганический фосфор чаще всего лимитирует первичную продукцию в пресных водах (Шиндлер, 1977; Хеки и Килхэм, 1988; Хадсон и др., 2000). Хотя концентрация Р в здоровых растениях невелика (0,1–0,8% сухой массы), он выполняет ключевые функции: входит в состав АТФ, АДФ, нуклеиновых кислот, коферментов, фосфолипидов и участвует в фосфорилировании сахаров (Рэйвен и др., 1981).
Многие исследования водотоков подтверждают, что фосфор ограничивает рост донных водорослей (Стокнер и Шортрид, 1978; Элвуд и др., 1981; Петерсон и др., 1983), однако в некоторых системах лимитирующим фактором выступает азот (Гримм и Фишер, 1986; Хилл и Найт, 1988) или оба элемента одновременно (Роземонд и др., 1993; Танк и Доддс, 2003). В данной главе рассматриваются три аспекта использования фосфора донными водорослями: (1) оценка ограниченности потребления Р, (2) измерение скорости поглощения Р, (3) определение скорости высвобождения Р (оборот). Следует отметить, что речь идёт только о неорганическом фосфоре, тогда как роль растворённого органического фосфора (ДОФ) остаётся малоизученной (Хванг и др., 1998; Пант и др., 2002). Термин «донные водоросли» означает водоросли, прикреплённые к погружённым субстратам, входящие в состав сложного сообщества — перифитона, ауфвухса или биоплёнки (включающей бактерии, грибы и мейофауну в полисахаридном матриксе; Лок и др., 1984).
А. Оценка степени ограниченности фосфора. Ограниченность биогенами у водорослей оценивают несколькими методами: по элементному составу биомассы, биоанализу обогащения, ферментативной активности и физиологическим реакциям. Элементный состав отражает дефицит, поскольку соотношения C:N:P в водорослях варьируют в зависимости от доступности питательных веществ (Холл и др., 2005). Эти пропорции влияют на трофические взаимодействия, динамику популяций, структуру сообщества и круговорот веществ (Хиллебранд и Келерт, 2001; Фрост и др., 2002). Согласно Калерту (1998), оптимальное молярное соотношение C:N:P для пресноводных донных водорослей составляет 158:18:1. Это отличается от редфилдовского соотношения 106:16:1 (Редфилд, 1958) для фитопланктона и 119:17:1 для морских донных микроводорослей (Хиллебранд и Соммер, 1999). Более высокое отношение C:P у донных форм объясняется присутствием богатого углеродом детрита, макроводорослей и слизистого матрикса. При дефиците фосфора соотношения C:P и N:P в тканях возрастают, так как клетки эффективнее используют доступный Р.
Для пресноводного планктона значения C:P >129 и N:P >22 (относительно соотношений Редфилда) указывают на умеренный дефицит фосфора (Хеки и др., 1993). Для донных водорослей пороги выше: дефицит предполагается при C:P >369 и N:P >32 (Калерт, 1998). Биоанализы обогащения заключаются в добавлении биогенов через диффузионные субстраты, порошкообразные удобрения или инъекции растворённых веществ. Эффект оценивают по изменению биомассы или первичной продукции по сравнению с контролем (главы 10, 28, 32). Надёжным ферментативным маркером дефицита фосфора служит активность фосфатазы (РА). Этот фермент гидролизует сложноэфирные связи, высвобождая ортофосфат (PO₄) из органических соединений. Чаще всего определяют щелочную фосфатазу, гидролизующую фосфомоноэфиры; также существуют фосфодиэстераза и АТФ-аза. Снижение концентрации неорганического фосфора ведёт к повышению активности РА (Хили, 1973; Ветцель, 1981; Эспеланд и др., 2002).
По данным Хили и Хендзела (1979), уровни фосфатазы выше 0,003 ммоль/мг хлорофилла а⁻¹ ч⁻¹ свидетельствуют об умеренном дефиците фосфора, а выше 0,005 — о серьёзном. Однако в дистрофических системах, богатых органическим фосфором, РА может давать смещённые результаты (Пант и др., 2002). Применение флуоресценции, меченной ферментами (ELF), позволяет определять дефицит фосфора на видовом уровне. Ренгефорс и др. (2001) использовали ELF для дифференциации РА у фитопланктона: флуоресцентный продукт образуется в месте ферментативной активности (на поверхности клетки). Метод также адаптирован для перифитонных биоплёнок, чтобы различать общую РА сообщества и активность отдельных бактерий (Эспеланд и др., 2002).
B. Показатели поглощения фосфора. Зависимость между концентрацией биогена в воде и скоростью его поглощения водорослями описывается гиперболической функцией Михаэлиса-Ментен:
V = Vm × (S / (Ks + S)) (уравнение 9.1),
где V — скорость усвоения, Vm — максимальная скорость, S — концентрация биогена, Ks — константа полунасыщения (концентрация, при которой скорость равна половине Vm). Из этого соотношения следуют два важных вывода. Во-первых, скорость поглощения растёт с увеличением концентрации фосфора. Насыщение может наступать при очень низких уровнях: <1 мкг/л для отдельных диатомей (Босуэлл, 1989) и <10 мкг/л для водотоков (Малхолланд и др., 1990). Во-вторых, значение Ks служит мерой сродства клетки к биогену: чем ниже Ks, тем выше сродство, что даёт преимущество при дефиците. Обычно Ks относительно стабильна, а при недостатке фосфора водоросли увеличивают Vm (Дарли, 1982; Ломан и Приску, 1992). Однако описаны многофазные системы поглощения Р (Ривкин и Свифт, 1982; Янссон, 1993), что указывает на неполную статичность Ks. Повышение Vm энергетически выгоднее, чем изменение Ks, так как требует лишь активации или синтеза большего количества существующего фермента. Длительное воздействие повышенных уровней биогенов может изменить видовой состав сообщества, увеличив долю видов с высокими Vm или иными Ks. Важно различать скорость поглощения и скорость роста, лимитированную биогенами; для описания роста используют модели Моно (внешняя концентрация) или Друпа (внутриклеточный пул; см. Друп, 1974; Ри, 1978; Борхардт, 1996).
C. Скорость обмена фосфора. Оборот фосфора отражает внутреннее перераспределение элемента в сообществе водорослей. После поглощения Р может включаться в структурные компоненты, сохраняться в лабильной форме или выделяться обратно. Клетки, испытывающие дефицит фосфора, с меньшей вероятностью высвобождают поглощённый Р, чем насыщенные (см. Сембелла и др., 1984; Борхардт и др., 1994). Следовательно, скорость оборота (потери фосфора клетками относительно общего содержания Р) у ограниченных клеток ниже, чем у насыщенных, при сходной метаболической активности и нагрузке пастбищ (Стейнман и др., 1995). Один из методов измерения оборота — мечение клеток радиоактивным фосфором (³²P или ³³P) в лаборатории, последующее возвращение водорослей в естественную среду и наблюдение за снижением радиоактивности. Получаемый показатель является кажущимся, так как оценивается по всей матрице перифитона, а не по отдельным клеткам.
D. Обзор главы. В данной главе сравнивается ограничение и поглощение фосфора донными водорослями из водотоков с низким и высоким содержанием Р. Теоретически, водоросли из условий низкого Р должны адаптироваться и проявлять следующие особенности (рис. 9.2): меньшую биомассу, более высокое соотношение C:P, повышенную активность фосфатазы, более низкие значения Ks, меньшую общую скорость поглощения (масса/время) и более низкую кажущуюся скорость оборота (большее удержание) по сравнению с сообществами из водотоков с высоким содержанием фосфора.
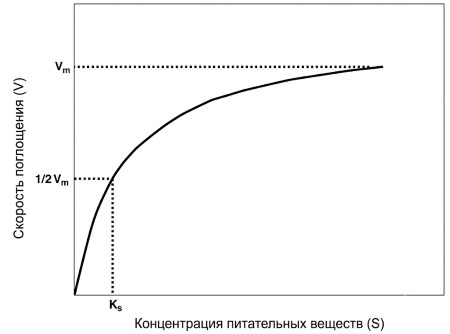
Рисунок 9.1. Зависимость между концентрацией биогена (S) и скоростью его усвоения (V). Vm — максимальная скорость усвоения; Ks — константа полунасыщения, т.е. концентрация, при которой скорость усвоения составляет половину от Vm.
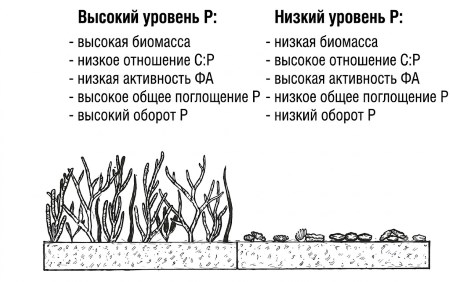
Рисунок 9.2. Гипотетические характеристики сообществ прикреплённых водорослей, подверженных воздействию высокого (слева) и низкого (справа) содержания фосфора: биомасса, соотношение C:P, активность фосфатазы, значения Ks, общая скорость поглощения и кажущаяся скорость оборота.
Сведения об авторах и источниках:
Авторы: Ф. Ричард Хауэр и Гэри А. Ламберти
Источник: Методы в экологии ручьев
Данные публикации будут полезны студентам экологических и географических специальностей, начинающим специалистам в области гидрологии, геоморфологии и управления водными ресурсами, а также всем, кто интересуется динамикой речных экосистем и вопросами охраны природных ландшафтов.
Дата добавления: 2026-05-20; просмотров: 268;
Поиск по сайту
Узнать еще
- Активные методы обучения в психологии: теория и практика применения
- Атмосферные процессы в смежных районах северного полушария
- Безопасность работы и методы установки подшипников
- Биоконтроль вредителей: методы, средства и роль биоразнообразия почвы
- Биология как наука: предмет изучения, методы и основные направления
- Блуждающие токи рельсовых сетей: природа, опасность электрокоррозии и методы защиты
- Виды потерь в волоконно-оптических системах связи и методы их минимизации
- Влияние неблагоприятных погодных условий на работу контактной сети и методы борьбы с ними

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
