Классификация взаимоотношений
Взаимоотношения организмов разнообразны (табл. 8). Они разделяются на горизонтальные – между организмами одного трофического уровня (как внутри вида, так и между видами) и ветрикальные – между организмами разных трофических уровней. Взаимоотношения первого рода, как правило, носят характер конкуренции, но могут на некоторых этапах жизни организмов быть мутуализмом (т.е. взаимопомощью). Взаимоотношения второго рода более разнообразны: «растение – фитофаг», «хищник – жертва» (иногда эти два типа взаимоотношений объединяют, так как растения по существу тоже жертвы), «паразит – хозяин», мутуализм, комменсализм, аменсализм.
Таблица 8 Типы взаимоотношений видов в экосистеме
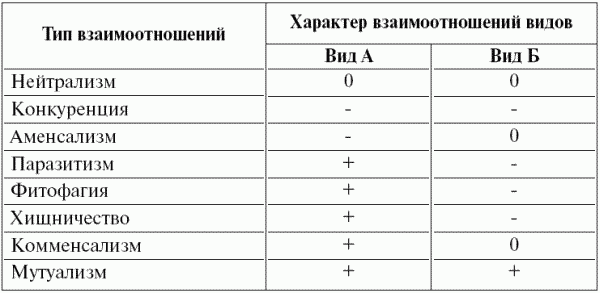
Примечание. Использованы следующие обозначения: 0 – отсутствие взаимоотношений, плюс – положительное влияние, минус – отрицательное влияние.
Кроме материальных взаимоотношений (конкуренции за ресурсы или передачи вещества и энергии при хищничестве или паразитизме), возможны сигнальные (информационные) взаимоотношения. Однако эти сигнальные взаимоотношения тесно переплетены с материальными и лишь корректируют распределение ресурсов между особями или передачу вещества и энергии с одного трофического уровня на другой.
Разделение взаимоотношений в естественных экосистемах по «полезности» и «вредности» некорректно: любые взаимоотношения помогают поддерживать экологическое равновесие и в конечном итоге являются «полезными» для всех видов, которые входят в состав экосистемы. О полезности и вредности отношений между организмами можно говорить только в том случае, если в эти отношения вмешивается человек.
Понятие «экологическое равновесие» подвергается жесткой критике некоторых экологов, которые считают, что этот феномен не существует в природе, а само понятие уходит корнями в представления Карла Линнея о божественной гармонии природы (см. 1.1). Оппоненты понятия «экологическое равновесие» считают, что любые взаимоотношения организмов и любая экосистема как сумма этих взаимоотношений всегда динамичны, так как испытывают влияние множества факторов.
Однако мы полагаем, что понятие экологического равновесия, если оно не абсолютизируется, а рассматривается как некоторая условность (подобная «идеальному газу» или «ускорению» под действием силы земного притяжения), полезно. Системы «хищник – жертва», «паразит – хозяин» и более сложные элементы экосистемы, такие, как пищевые цепи, в состоянии экологического равновесия характеризуются соотношением численности и биомассы взаимодействующих организмов, которые колеблются вокруг некоторой средней величины, соответствующей «экологическому равновесию». Это понятие необходимо при разработке системы рационального использования и охраны природы. Возможно, что более точно этот феномен отражают термины «динамическое равновесие» или «подвижное равновесие».
Контрольные вопросы
1. Назовите основные типы вертикальных и горизонтальных отношений между видами.
2. Почему некорректно разделять взаимоотношения популяций в природе на «полезные» и «вредные»?
3. Что понимается под экологическим равновесием?
Конкуренция
Конкуренция – это соревнование организмов одного трофического уровня (между растениями, между фитофагами, между хищниками и т.д.) за потребление ресурса, имеющегося в ограниченном количестве. Д.Тилман (Tilman, 1982, 1983) подчеркивает, что особую роль играет конкуренция за потребление ресурсов в критические периоды их дефицита (например между растениями за воду в период засухи или хищниками за жертвы в неблагоприятный год). Конкуренция играет большую роль в определении видового состава экосистем.
Принципиальных различий у межвидовой и внутривидовой (внутрипопуляционной) конкуренции нет. Возможны как случаи, когда внутривидовая конкуренция является более острой, чем межвидовая, так и наоборот. При этом интенсивность конкуренции внутри популяции и между популяциями может меняться в различных условиях. Если условия неблагоприятны для одного из видов, то конкуренция между его особями может усиливаться. В этом случае он может быть вытеснен (или чаще – потеснен) видом, для которого эти условия оказались более подходящими.
Однако в многовидовых сообществах пар «дуэлянтов» чаще всего не образуется, и конкуренция носит характер диффузной: много видов одновременно конкурируют за один или несколько факторов среды. «Дуэлянтами» могут быть лишь массовые виды растений, которые делят один и тот же ресурс (например, деревья – липа и дуб, сосна и ель и т.д.).
У растений возможна конкуренция за свет, за ресурсы почвы и за опылителей. На почвах, богатых ресурсами минерального питания и влагой, формируются густые сомкнутые растительные сообщества, где лимитирующим фактором, за который конкурируют растения, является свет. При дефиците в почве влаги или элементов минерального питания полог растений бывает разомкнутым и они не конкурируют за свет, а соревнуются за потребление почвенных ресурсов (рис. 19).

Рис. 19. Основной комплексный градиент конкуренции растений (по Tilman, 1988).
При конкуренции за опылителей побеждает тот вид, который более привлекателен для насекомого. Так одним из факторов быстрого распространения в Европе гималайского заносного вида недотрога железконосная (Impatiens glandulifera) является то, что он продуцирует больше нектара, чем его конкуренты в тех же влажных местообитаниях – чистец болотный, дербенник иволистный. Кроме того, нектар недотроги слаще (Chittka, Schurkens, 2001).
У животных конкуренция происходит за ресурсы пищи, например травоядные конкурируют за фитомассу При этом конкурентами крупных копытных могут быть насекомые, подобные саранче, или мышевидные грызуны, способные в годы массового размножения уничтожить большую часть травостоя. Хищники конкурируют за жертвы.
Поскольку количество пищи зависит не только от экологических условий, но и от площади, где воспроизводится ресурс, конкуренция за пищу может перерастать в конкуренцию за занимаемое пространство, т.е. быть не только эксплуатационной, но и интерференционной (см. 8.2). Снижение конкуренции в этом случае возможно при разделении территории на «охотничьи наделы» или на «загоны для выпаса». К примеру, косяки – семейные группы башкирской лошади, в поведении которых сохранились черты их диких предков, рассредоточиваются по «загонам для выпаса», что снижает конкуренцию за фитомассу и способствует равномерному использованию травостоев.
По этой причине лошади – это идеальные фитофаги для особо охраняемых территорий в степной зоне.
Большую роль в разделе территории играют сигнальные взаимоотношения (см. 8.8).
Как и в отношениях между особями одной популяции, конкуренция между видами (их популяциями) может быть симметричной или асимметричной. При этом ситуация, когда условия среды одинаково благоприятны для конкурирующих видов, встречается довольно редко, и потому отношения асимметричной конкуренции возникают чаще, чем симметричной.
При флюктуирующих ресурсах, что обычно в природе (увлажнение или элементы минерального питания для растений, первичная биологическая продукция для разных видов фитофагов, плотность популяций жертв для хищников), поочередно получают преимущества разные конкурирующие виды. Это также ведет не к конкурентному исключению более слабого, а к сосуществованию видов, которые поочередно попадают в более выгодную и менее выгодную ситуацию. При этом ухудшение условий среды виды могут переживать при снижении уровня метаболизма или даже перехода в состояние покоя.
Кроме того, на исход конкуренции часто влияет то, какой из видов первым начал заселять экотоп (принцип лотереи, см. 12.8). Это особенно характерно для маловидовых сообществ водных растений, где более слабый вид может удерживать занятое место, если ему повезло и он его занял первым. Впрочем, лотерея может влиять и на состав сообществ с большим числом претендентов на свободное место. В тропическом лесу на одном гектаре может быть до 150 видов деревьев, и потому занять место выпавшей особи могут представители разных видов.
Влияет на исход конкуренции и то, что победить в конкурентной борьбе больше шансов имеет популяция, в составе которой больше особей и которая, соответственно, будет более активно воспроизводить «свою армию» (так называемый масс‑эффект).
Наконец, конкуренция между видами протекает на фоне отношений с организмами других трофических уровней (хищниками и паразитами). Это также влияет на исход конкуренции, так как более привлекательный как пищевой ресурс вид имеет меньше шансов победить в конкуренции. В итоге в естественных экосистемах виды сосуществуют даже при наличии асимметричной конкуренции, которая должна была бы привести к вытеснению одного из видов. Конкурентное исключение чаще всего наблюдается только в искусственных условиях «микрокосма», когда два конкурирующих вида изолированы и помещены в условия стабильной среды (например в смешанном посеве двух культурных растений с разными конкурентными возможностями).
В естественных экосистемах существуют и специальные механизмы, которые снижают конкуренцию. Главный механизм – дифференциация экологических ниш (см. 9), при которой разные организмы используют разные ресурсы.
Контрольные вопросы
1. Дайте определение конкуренции.
2. Имеются ли принципиальные отличия внутривидовой конкуренции от межвидовой?
3. Какая конкуренция называется асимметричной?
4. Что такое диффузная конкуренция?
5. Приведите примеры конкуренции животных за разные ресурсы?
6. За какие ресурсы среды конкурируют растения?
7. Чем отличаются эксплуатационная и интерференционная виды конкуренции?
8. Как влияет на конкуренцию «принцип лотереи»?
9. Как влияет на конкуренцию «масс‑эффект»?
10. Какое значение для конкуренции имеют флюктуации количества ресурсов?
Дата добавления: 2020-10-01; просмотров: 1058;
Поиск по сайту
Узнать еще
- II Классификация САSЕ-средств
- II. Классификация документов
- II.4. Классификация нефтей и газов по их химическим и физическим свойствам
- III. Классификация методов воспитания.
- III.1.3. ПРИЧИНЫ НАРУШЕНИЙ СЛУХА. ПСИХОЛОГО-ПЕДАГОГИЧЕСКАЯ КЛАССИФИКАЦИЯ НАРУШЕНИЙ СЛУХОВОЙ ФУНКЦИИ У ДЕТЕЙ
- III.2.3. ПРИЧИНЫ НАРУШЕНИЙ ЗРЕНИЯ. КЛАССИФИКАЦИЯ НАРУШЕНИЙ ЗРИТЕЛЬНОЙ ФУНКЦИИ У ДЕТЕЙ
- VI.2. Классификация месторождений нефти и газа
- VI.III. VI. Генетическая классификация складок.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
