Общая характеристика
Тело гриба представляет собой мицелий, состоящий из тонких нитей - гиф. Мицелий имеет тесную связь с субстратом, что обусловлено осмотическим поглощением питательных веществ. У высших грибов мицелий разделен на отдельные клетки перегородками - септами, т.е. у них септированнный (клеточный) мицелий. Низшие грибы имеют неклеточное строение мицелия, так как его гифы не разделены на перегородки, а представляют собой как бы одну разветвленную клетку со множеством ядер.
Грибы обособлены по своей морфофизиологической организации от остального мира живых существ. Их нельзя отнести ни к растениям, ни к животным. Существуют две теории происхождения грибов: животная и растительная, поскольку клетки грибов имеют признаки как животной, так и растительной клетки (табл. 5.2).
Теория растительного происхождения грибов предполагает их происхождение от зеленых водорослей, из чего следует, что грибы прежде всего явно регрессивная группа растений, утративших хлоропласты.
Теория животного происхождения основывается на том, что грибы изначально являются бесхлорофильными организмами, т.е. происходят от простейших гетеротрофных организмов, а не от водорослей. Эта теория предпочтительнее, поскольку бесхлорофильные водоросли, относимые к зеленым, в качестве запасного продукта накапливают крахмал, в то время как у грибов крахмала нет.
Таблица 5.2.Особенности строения грибной клетки

Грибы - это гетеротрофы. Как и бактериям, им свойственно внеклеточное переваривание, осуществляемое за счет выделения во внешнюю среду ферментов. Всасывание расщепленных питательных веществ происходит осмотически, всей поверхностью тела. Клетки мицелия в качестве запасных питательных веществ откладывают углеводы в виде гликогена, жиры в виде капель липидов, а в вакуолях - белки.
Грибы могут быть паразитами, сапрофитами и симбионтами. Среди паразитов выделяют облигатные и факультативные. Облигатные паразиты узко специализированы и имеют небольшой круг хозяев (мучнеросные, ржавчинные и головневые). Циклы развития гриба строго согласованы с продолжительностью жизни хозяина. При этом на гифах гриба образуются присоски-гаустории, которые внедряются в клетку для поглощения из нее пищи. Факультативные паразиты лишь временно способны к паразитизму. Они не образуют гаусторий, а с помощью ферментов разрушают ткань растения, поглощают содержимое клеток и вызывают гибель хозяина. В дальнейшем они питаются сапрофитно мертвыми остатками (например, картофельный гриб фитофтора, поражающий многие растения семейства Пасленовые).
Грибы способны вступать в симбиоз с высшими растениями, образуя микоризу (грибокорень). Грибы используют углеводы, синтезируемые растением, и добывают для него (за счет минерализации органических соединений) различные соединения с азотом, фосфором, вырабатывают активаторы роста и витаминоподобные вещества.
Размножатьсягрибы могут вегетативно, бесполым и половым путем.
Вегетативное размножение может происходить частями мицелия (почти у всех грибов), почкованием (дрожжи). Бесполое размножение происходит за счет образования зооспор, спорангиоспор и конидий.
Зооспоры образуются у грибов, ведущих водный образ жизни (хи- тридиомицеты, оомицеты). Их подвижность обеспечивают жгутики (их 1 или 2). Образуются внутри одноклеточных зооспорангиев и при созревании выходят в воду. Покрываются оболочкой и прорастают в новую особь.
Спорангиеспоры образуются эндогенно - внутри одноклеточных спорангиев, возникающих на гифах-спорангиеносцах. В одном спорангии может быть до 10 тыс. спор, которые при созревании выходят из спорангия и распространяются ветром на значительные расстояния. Попав в благоприятные условия, спора прорастает в новый мицелий (например, у мукора).
Конидии образуются экзогенно на особых гифах - конидиеносцах. Конидии образуют цепочки, отчленяются и в благоприятной среде прорастают в новый мицелий (например, у пеницилла).
Половое размножение у низших грибов происходит:
• при слиянии гамет - гаметогамия (изогамия, гетерогамия и оогамия);
• при слиянии двух многоядерных специализированных половых органов (гаметангиев) - зигогамия.
Половое размножение у высших грибов:
• гаметангиогамия; архикарп - женский гаметангий, антеридий - мужской (у сумчатых грибов);
• соматогамия - слияние гаплоидных соматических клеток гетероталличных гиф (+ и - физиологически различных гиф), например у высших базидиальных грибов.
Половой процесс всегда завершается образованием диплоидной зиготы, ее мейотическим делением и спороношением.
К низшим грибам относится отдел зигомикоты, к высшим - отделы: сумчатые, базидиомикоты, несовершенные.
ОТДЕЛ ЗИГОМИКОТЫ(ZYGOMYCOTA)
Почти все представители этого класса ведут наземный образ жизни. Среди них есть как сапрофиты, так и паразиты высших грибов, насекомых, животных и человека. Мицелий в основном неклеточный, клеточные стенки содержат хитин. Наиболее известен род мукор (головчатая плесень) - Mucor.
Мукор широко распространен в природе как белая плесень (рис. 5.15). Сапрофит по способу питания; развивается на почве, пищевых продуктах. Гифы мицелия представляют собой вытянутую разросшуюся гигантскую клетку с множеством ядер (неклеточное строение). Ядра - с гаплоидным набором хромосом (n). На мицелии развиваются многочисленные вертикальные спорангиеносцы с буро-черными спорангиями. В результате митоза содержимое спорангия распадается на множество спор (до 10 тыс.). После созревания оболочка спорангия лопается, и споры рассеиваются, прорастая в новые особи. Размножение может быть бесполым (спорами), вегетативным (частями мицелия), редко - половым (зигогамия).
При зигогамии (рис. 5.16) физиологически различные гифы - гетероталличные, условно обозначающиеся как + и -, начинают расти навстречу друг к другу. На концах гиф образуются гаметангии, от- деляющиеся перегородками от остальной гифы. Далее происходит гаметангиогамия, состоящая в слиянии 2 специализированных половых структур (гаметангиев), не дифференцированных на гаметы, и образуется зигота с множеством диплоидных ядер. Зигота покрывается толстой бурой оболочкой. После периода покоя ядра претер- певают мейоз, а зигота прорастает в зародышевый спорангий. В него переходят гаплоидные ядра + и-, образовавшиеся после мейоза. В спорангии формируются споры, после их созревания спорангий вскрывается, споры рассеиваются и прорастают в новые мицелии (+ и -).
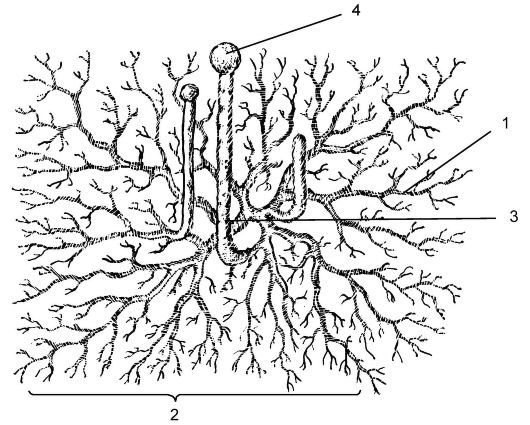
Рис. 5.15.Строение мукора (Мuсоr mucedo): 1 - гифа; 2 - мицелий; 3 - спорангиеносец; 4 - спорангий со спорами
Некоторые мукоровые грибы вызывают микоз (мукоромикоз) легких (ложный туберкулез), головного мозга и других органов че ловека, а также сельскохозяйственных растений. Многие виды рода обладают высокой ферментативной активностью, что используется при получении «соевого сыра» из семян сои, спирта из клубней картофеля и т.д.
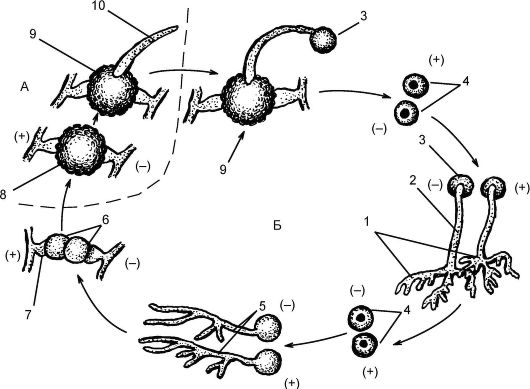
Рис. 5.16.Жизненный цикл мукора (Мuсоr): А - гаплоидная фаза; Б - диплоидная фаза: 1 - два гетероталличных (противоположных по физиологическому знаку) мицелия; 2 - спорангиеносец; 3 - спорангий; 4 - споры; 5 - прорастание спор; 6 - гаметангий; 7 - подвесок; 8 - зигоспора; 9 - прорастающая зигоспора; 10 - прорастающий мицелий
ОТДЕЛ СУМЧАТЫЕ ГРИБЫ, ИЛИ АСКОМИКОТЫ(ASCOMYCOTA)
Это один из наиболее обширных классов грибов, включающий более 30 тыс. видов. К этому классу относятся дрожжи, представ- ленные одиночными почкующимися клетками, и грибы с крупными плодовыми телами, например сморчки и строчки. Аскомикоты широко распространены в природе во всех природных зонах. По способу питания это сапрофиты. Мицелий сумчатых грибов является септированным, т.е. разделен на клетки (с гаплоидным набором хромосом). Характерный признак аскомикот - наличие сумок (аск), образующихся в результате полового процесса. Сумки - замкнутые структуры, содержащие определенное количество аскоспор (споры полового размножения) и образующиеся в результате мейоза.
У многих аскомикот сумки образуются в плодовых телах (подкласс Плодосумчатые). Различают 3 типа плодовых тел: клейстотеций, перитеций и апотеций. У других представителей сумки лежат открыто на мицелии (подкласс Голосумчатые).
Большая роль в цикле развития принадлежит и бесполому размножению. Споры бесполого размножения - конидии - образуются в результате митоза на мицелии с гаплоидными ядрами (n) или кони- диеносцах различного строения.
Подкласс Голосумчатые(Hemiascomycetidae)
Наиболее распространенным и имеющим практическое значение является род Дрожжи (Saccharomyces). Дрожжи представлены единичными, овальными клетками (рис. 5.17). Для дрожжей харак- терно вегетативное размножение, осуществляющееся почкованием; для этого им необходимы питательная среда, наличие в ней сахара и определенная температура. При неблагоприятных условиях происходит половой процесс; при слиянии 2 гаплоидных дочерних клеток (хологамия) образуется зигота, превращающаяся в сумку. В результате мейоза в сумке образуются четыре споры (аскоспоры), прорастающие в новые дрожжевые клетки.
Пекарские дрожжи (Saccharomyces cerevisiae) объединяют многие выведенные в культуре дрожжи: спиртовые, пивные, винные, хлебопекарные. Все эти дрожжи разлагают сахар на этиловый спирт и СО2. Так, при добавлении дрожжей в тесто они начинают разлагать имеющуюся там глюкозу, образующуюся из крахмала. При этом выделяется СО2, обеспечивающий тесту пористость и увеличение объема. При выпечке этанол и СО2 улетучиваются.
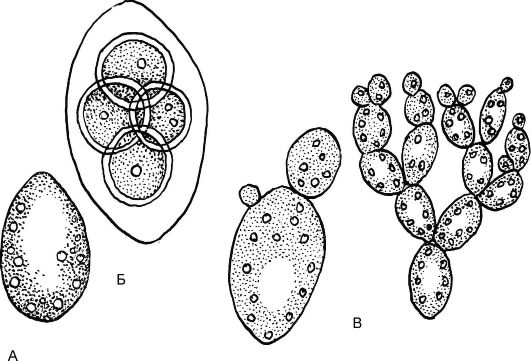
Рис. 5.17.Дрожжи пивные (Saccharomyces cerevisiae): А - одноклеточный таллом; Б - сумка с аскоспорами; В – почкование
Дрожжи являются ценным пищевым и кормовым продуктом. Содержат до 50% белка, а также жиры и углеводы. Синтезируют в больших количествах витамины, особенно В2. Их используют при лечении малокровия, а также как источник белка при добавлении в кормовые продукты в животноводстве и птицеводстве.
Подкласс Плодосумчатые(Carpoascomycetidae)
Представители этого подкласса характеризуются наличием плодовых тел, в которых находятся сумки. Плодовые тела образуются за счет плотного сплетения гаплоидных и дикарионных (двуядерных) гиф, называемых также аскогенными. Плодовые тела (аскокарпы) бывают 3 типов: замкнутые (закрытые) - клейстотеции, полузамкнутые - перитеции, незамкнутые (открытые) - апотеции.
К грибам-паразитам относится род Спорынья (Claviceps). Большинство представителей рода паразитируют на злаках - культивируемых (рожь, пшеница, ячмень) и дикорастущих (тимофеевка, пырей, костер). Самый распространенный вид - Спорынья пурпурная (Claviceps purpurea).
Цикл развития спорыньи протекает со сменой ядерных фаз (рис. 5.18). Так, осенью на злаковых растениях образуются склеро- ции - темно-фиолетовые снаружи и белые внутри рожки, представляющие собой мицелий гриба (обезвоженные гифы) в стадии покоя. На зиму склероции выпадают из злаков на почву и зимуют в ней. Весной на почве склероции прорастают, образуя нитевидные выросты, увенчанные головками - стромами. В этих стромах в результате полового процесса образуются плодовые тела - перитеции, заполнен- ные длинными цилиндрическими сумками (асками), содержащими нитевидные аскоспоры - споры полового размножения (рис. 5.19). Созревание спор идет в результате мейоза во время цветения злака. Споры активно выбрасываются с помощью ветра, попадают на рыльце цветущего злака и прорастают. Образующийся мицелий проникает в завязь пестика и разрушает ее. На концах гиф мицелия в результате митоза отшнуровываются конидии - споры бесполого размножения, т.е. наступает конидиальное спороношение. При этом гифы гриба выделяют капельки сладкой жидкости - «медвяную росу». Насекомые переносят конидии на цветки соседних растений и заражают их.
Склероции содержат высокотоксичные алкалоиды, вызывающие отравления. Такие алкалоиды, как эрготамин, эргометрин, широко применяют в современной медицине для лечения сердечнососудистых, нервных и других заболеваний. Особенно эффективны они в акушерско-гинекологической практике.
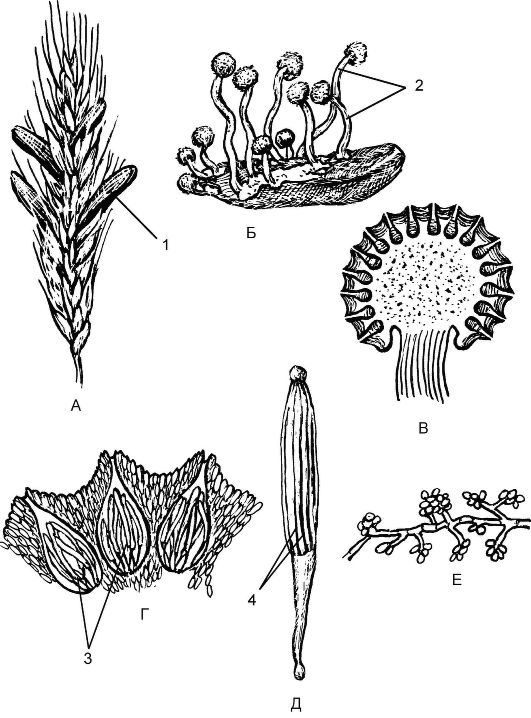
Рис. 5.18.Спорынья пурпурная (Claviceps рurpurеа): А - колос ржи со склероциями (1); Б - стромы (2), выросшие на перезимовавшем склероции; В - продольный срез через строму с перитециями; Г - продольный срез через перитеции (3) с сумками; Д - сумка с нитевидными аскоспорами (4); Е - конидиальное спороношение
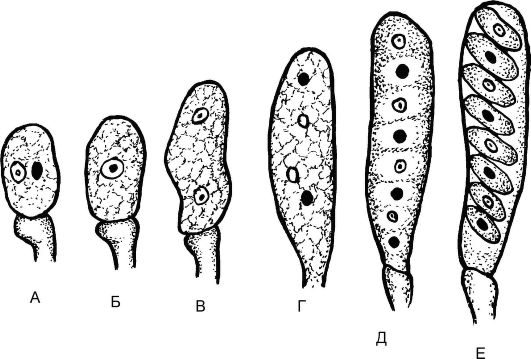
Рис. 5.19.Развитие сумки с аскоспорами: А, Б - образование зиготы на верхушке аскогенной гифы; В-Е - мейоз и развитие сумки с аскоспорами
Незамкнутые плодовые тела - апотеции - встречаются у таких представителей, как сморчки (Morchella), строчки (Gyromitra). Это открытое плодовое тело обычно блюдцевидной, бокаловидной формы размером от 0,1 до 10 см, разнообразной окраски - от ярко-оранжевой или красной до коричневой и черной. Верхний слой (гимений) содержит множество сумок. Плодовые тела грибов из этой группы состоят из стерильной ножки и складчатой или лопастной шапки (рис. 5.20).
Сморчки и строчки - съедобные грибы, но при употреблении в пищу строчки предварительно необходимо прокипятить, а воду слить.
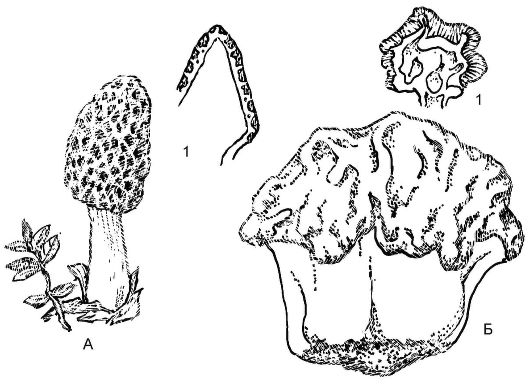
Рис. 5.20.Аскомикоты - внешний вид и плодовые тела сморчков и строчков:
А - сморчок конический (Моrchella coinca); Б - строчок обыкновенный (Gyromitra exculenta); 1 - разрезы плодовых тел
ОТДЕЛ БАЗИДИОМИКОТЫ(BASIDIMYCОTА)
В этом классе объединены почти все группы шляпочных грибов, насчитывающих около 30 тыс. видов. Вегетативное тело представлено членистым мицелием, состоящим из членистых гиф.
Размножение:вегетативное (осуществляется частями мицелия), бесполое (конидиями) и половое.
При половом процессе специальных органов полового размножения не образуется. Половой процесс протекает в форме соматогамии (рис. 5.21). Из прорастающей гаплоидной базидиоспоры развивается первичный мицелий, который затем превращается в членистый. Каждый членик одноядерный. Вскоре происходит гологамия - слияние конечных клеток гиф. Однако слияние содержимого члеников не сопровождается слиянием ядер. Образуются дикарионы, которые затем синхронно делятся. Так формируется вторичный дикарионный мицелий.

Рис. 5.21.Развитие базидиального гриба. Схема цикла развития: А - схема цикла развития: 1 - базидия; 2 - базидиоспора; 3 - первичный мицелий; 4 - дикарионный мицелий; 5 - плодовое тело из дикарионного мицелия; Б - развитие базидии с базидиальными спорами
На дикарионном мицелии образуется плодовое тело, которое состоит из пенька и шляпки. Гимениальный слой шляпки может быть пластинчатым или трубчатым. В гимениальном слое на концах дикарионных гиф из 2 ядерных клеток образуются базидии. По своему развитию базидии гомологичны сумкам. В базидии завершается половой процесс, т.е. сливаются ядра дикариона и образуется диплоидное ядро. Такая одноклеточная базидия называется холобазидией. Образовавшееся диплоидное ядро делится мейозом с образованием 4 гаплоидных ядер (см. рис. 5.19, А). К этому времени в верхней части базидии образуются четыре трубчатых выроста - стеригмы. В стеригмы перетекают образовавшиеся ядра и формируются 4 базидиоспоры: 2 условно со знаком - и 2 со знаком +. Поэтому первичные мицелии, вырастающие из них, будут гетероталличными. Базидии образуются непосредственно на гифах или в плодовых телах различной формы, но чаще состоящих из шляпки и ножки. В цикле развития чередуются 3 фазы: гаплоидная (короткая) - это базидиоспоры, дикарионная (длится основную часть жизни) - дикарионный мицелий и диплоидная (кратковременная) - молодая базидия до образования базидиоспор.
ОТДЕЛ ДЕЙТЕРОМИКОТЫ(DEUTEROMYCOTA), ИЛИ НЕСОВЕРШЕННЫЕ ГРИБЫ(FUNGI IMPERFECT!)
Дейтеромикоты, наряду с бизидиомикотами и аскомикотами, являются крупнейшей группой грибов, объединяющей 25-30 тыс. ви- дов. Эти грибы представляют собой бесполые формы (анаморфы), размножающиеся бесполым путем - конидиями. Жизненный цикл у них проходит в гаплоидной стадии без полового процесса. Вполне возможно, что дейтеромикоты - это наиболее специализированные линии эволюции грибов.
Дейтеромикоты широко распространены в природе и встречаются на различных субстратах, принимая вместе с другими грибами участие в разложении органических остатков и в почвообразовательном процессе. Многие грибы этого отдела паразитируют на высших растениях, вызывая серьезные болезни сельскохозяйственных культур. Аспергилл паразитический (Aspergillus parasiticus) выделяет аффлатоксины, очень опасные из-за своей канцерогенности. У части представителей этих родов известны также сумчатые стадии, в связи с чем их нередко рассматривают среди аскомикот. Пенициллы и аспергиллы широко используются в микробиологической промышленности для биотехнологического производства ряда органических кислот (лимонной, фумаровой, глюконовой и др.), ферментов (протеиназ, амилаз и др.) и антибиотиков (пенициллина, фумагиллина). Антибиотики подавляют рост других организмов (особенно бакте- рий), поэтому их применяют для лечения многих заболеваний

Большое медицинское значение имеет род Пенициллиум (Penicillium). Пеницилл имеет членистый мицелий зеленоватого цвета, состоящий из одноядерных члеников. Отходящие вверх гифыконидиеносцы ветвятся на верхнем конце на стеригмы. Последние по внешнему виду напоминают кисточку или кисть руки и заканчиваются цепочкой наружных спор - конидий (рис. 5.22). Конидии - это споры бесполого размножения, образующиеся за счет митоза.
Наблюдается и половой процесс, в результате которого непосредственно на мицелии образуются закрытые шаровидные плодовые тела ярко-желтого цвета - клейстотеции. Внутри клейстотециев формируются сумки с 8 аскоспорами. Созревшие аскоспоры выходят из сумок после разрыва клейстотеция.
Пеницилл (Penicillium), сапрофит по способу питания, поселяясь на пищевых продуктах и изделиях (ткани, кожа), вызывает их порчу. Пеницилл используют не только в медицинской практике, но и в пищевой промышленности для приготовления особых сортов сыра («рокфор»).
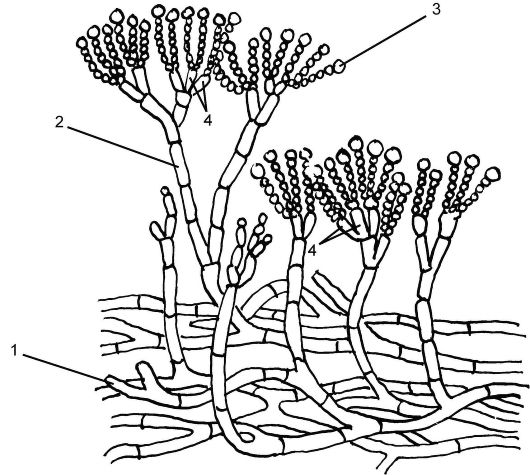
Рис. 5.22.Дейтеромикоты (Deuteromycota) пеницилл: 1 - мицелий; 2 - конидиеносец; 3 - конидии; 4 – стеригмы
Велико значение грибов в деятельности человека. Они участвуют в круговороте веществ в природе. Грибы, как и бактерии, минерализуют органические вещества и принимают участие в образовании гумуса. Их используют в пищевой промышленности для производства спирта, вина, пива, кваса, в хлебопечении, при получении белков и витаминов. Грибы образуют органически активные вещества - антибиотики, ферменты, органические кислоты и др.
Грибы могут вызывать коррозию металлов, разрушать кожу, бумагу, ткани. Многие грибы наносят существенный вред человеку, животным и растениям, вызывая ряд заболеваний (микозы, стригущий лишай, паршу), а также приводить к порче пищевых продуктов и тем самым служить причиной различных отравлений.
ОТДЕЛ ЛИШАЙНИКИ(LICHENES)
Это группа симбиотропных растений, состоящих из 2 компонентов - автотрофных водорослей и гетеротрофных грибов. Грибная основа лишайников формируется в основном сумчатыми грибами. Водорослевый компонент состоит из видов, относимых в большинстве случаев к представителям отделов зеленые и сине-зеленые водоросли. Выделенные из лишайника водоросли не отличаются от свободно живущих форм. Физиологически этот тип симбиоза основан на межклеточном обмене между водорослями и грибами. Гриб питается углеводами водоросли, а водоросли получают от грибов минеральные вещества. Однако симбиоз с грибами приводит к появлению нового биологического качества, которое выражается у лишайника в его способности размножаться как единый организм.
Вегетативное тело лишайников представлено слоевищем, имеющим различную окраску (серую, зеленоватую, буро-коричневую, желтую или почти черную). Морфологически различают 3 основных типа слоевища лишайников: накипной (корковый), листоватый и кустистый (рис. 5.23), однако существуют и переходные формы. Наиболее низкоорганизованные - накипные, или корковые, слоевища; они имеют вид порошковатых, зернистых, бугорчатых налетов, плотно срастающихся с субстратом и не отделяющихся от него без значительных повреждений.
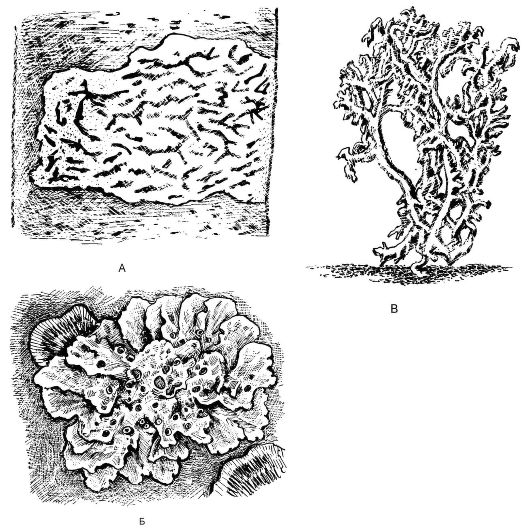
Рис. 5.23.Различные типы слоевищ лишайников: А - корковый (графис - Graphis scripta); Б - листоватый (ксантория - Xanthoria); В - кустистый (кладония - Cladonia)
Более высокоорганизованные лишайники имеют листоватое слоевище в форме пластинок, чешуек или розеток, прилепленных к по- чве или деревьям с помощью ризин - аналогов ризоидов, состоящих из пучков грибных гиф.
Наивысшей организации в своем строении достигают лишайники с кустистым типом слоевища, имеющие вид разветвленного кустика (12-15 см в высоту) и срастающиеся с субстратом только основанием.
По анатомическому строению лишайники бывают гомеомерными и гетеромерными (рис. 5.24). У более примитивных - гомеомерных - по всей толще слоевища равномерно расположены грибные гифы и водоросли. При гетеромерном строении на поперечном разрезе лишайника сверху можно видеть так называемую верхнюю кору. Она образована переплетающимися и тесно смыкающимися грибными гифами. Под корой грибные гифы лежат более рыхло, и между ними находятся клетки водоросли (гонидиальный слой). Внутри слоевища можно выделить сердцевину, состоящую из рыхлых грибных гиф и больших пустот, заполненных воздухом. Под ней размещается нижняя кора, которая по строению сходна с верхней. Через нее из сердцевины проходят отдельные гифы (ризины), закрепляя лишайник в субстрате.
Большинство лишайников легко переносят высыхание. Фотосинтез и питание у них в это время прекращаются, чем и объясняется их незначительный ежегодный прирост.
Размножениелишайников преимущественно вегетативное, осно- вано на способности лишайников регенерировать из отдельных участков. Оно осуществляется путем фрагментации (отделения участков слоевища) или с помощью обособленных групп клеток водорослей, окруженных гифами гриба и различных по своей форме, - соредий, изидий и лобул (рис. 5.25). Соредии - мельчайшие образования округлой формы, включающие одну или несколько клеток водоросли и окруженные грибными гифами. Изидии - бугорчатые палочковидные выросты на верхней поверхности слоевища.
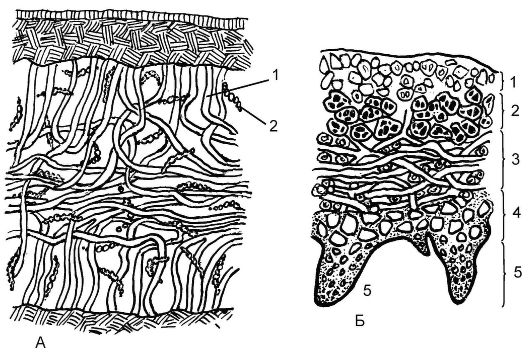
Рис. 5.24.Анатомическое строение слоевищ лишайников: А - разрез гомеомерного слоевища лишайника: 1 - гифы гриба; 2 - водорослевый компонент;
Б - разрез гетеромерного лишайника: 1 - верхний корковый слой; 2 - гонидиальный слой; 3 - средний слой с гифами гриба; 4 - нижний корковый слой; 5 - ризины
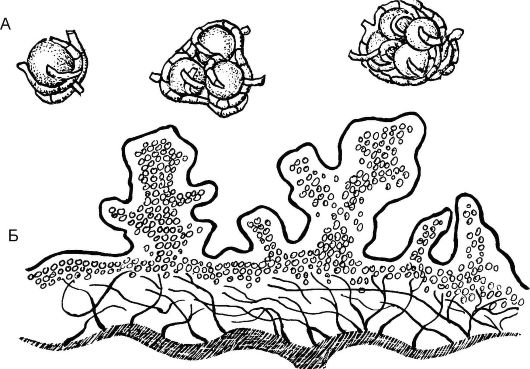
Рис. 5.25.Размножение лишайников: А - соредии; Б - изидии
Лобулы имеют вид маленьких чешуек, расположенных вертикально на поверхности слоевища или по его краям. Кроме того, наблюдается бесполое размножение с помощью спор, самостоятельно образующихся и у водорослей, и у грибов.
Половое размножение изучено недостаточно, но в общих чертах протекает так же, как у свободно живущих грибов.
Значение лишайников велико. Они разлагают и минерализуют ор- ганические вещества почвы. Являются первопроходцами - одними из первых заселяя скалы, они разрушают их поверхностный слой и, отмирая, образуют гумус, на котором расселяются другие растения. Лишайники являются индикаторами чистоты воздуха, так как не выносят даже малейших примесей сернистых газов. Из некоторых их видов получают краску и особое вещество - лакмус (для химической промышленности). В тундре и лесотундре лишайники (ягель) являются основным кормом для оленей. Встречаются съедобные лишайники также в полупустынных и пустынных районах Киргизии и Туркмении.
Дата добавления: 2016-05-30; просмотров: 13914;
Поиск по сайту
Узнать еще
- Cравнительная характеристика усилителей на БТ
- I. Загальна характеристика
- II. Монозы и их характеристика
- Nemathelmintes. Общая характеристика типа. Nematoda. Характеристика класса. Медицинское значение. Био- и геогельминты.
- Plahelmintes (Плоские черви).Общая характеристика типа. Морфология, систематика, основные представители, значение.
- Protozoa. Общая характеристика подцарства. Классификация. Представители. Медицинское значение.
- V. Механическая характеристика
- V. Упрощенная схема замещения трансформатора и внешняя характеристика.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
