Антигенраспознающие рецепторы В-лимфоцитов
Антигенраспознающие рецепторы В-лимфоцитов представляют собой мономерные формы иммуноглобулина М, встроенные в плазматическую мембрану В-лимфоцита и окруженные специальными вспомогательными трансмембранными белками – Iga и Igb, которые принимают участие в передаче сигнала от IgМ, присоединившего антиген, внутрь клетки. Кроме того, эти белки необходимы для нормального выхода мембранной формы IgМ в плазматическую мембрану: в отсутствии этих белков молекулы мембранного IgМ остаются внутри клетки.
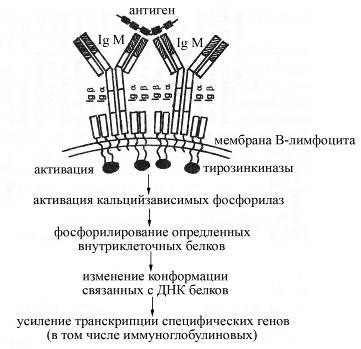
Рис. Схема строения антигенраспознающего рецептора В-лимфоцитов и механизма их активации после взаимодействия с антигеном. Антигенраспознающий рецептор В-лимфоцитов представлен мембранной формой IgМ, окруженного специальными вспомогательными белками – Igα и Igβ, принимающими участие в передачи сигнала от IgМ внутрь клетки
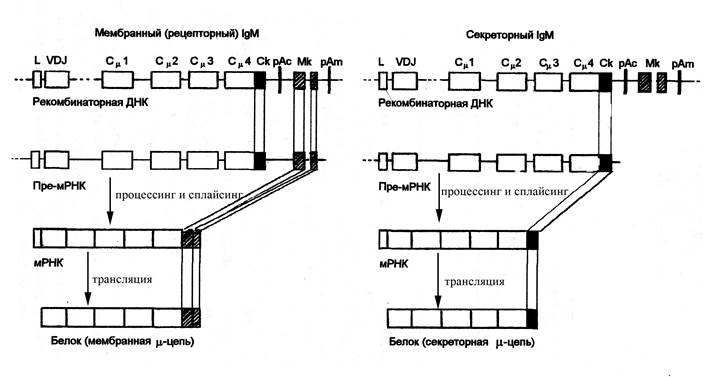 Мембранная форма иммуноглобулина М отличается от секреторной не только тем, что является мономерной, но и некоторыми особенностями строения самого мономера IgМ: в его состав входит гидрофобный трансмембранный участок и короткий гидрофильный цитоплазматический хвост. В то же время мономерная молекула секреторной формы иммуноглобулина М имеет на С-конце углеводные остатки, увеличивающие растворимость молекулы, и цистеиновый остаток, необходимый для полимеризации мономеров в пентамерную форму с помощью J-цепи. Молекула мембранного иммуноглобулина М кодируется теми же рекомбинантными иммуноглобулиновыми локусами, что и секреторного иммуноглобулина (χ- или l-локусом, кодирующим легкую цепь, и локусом тяжелых цепей). Некоторые же отличия в строении С-конца тяжелых цепей секреторного и мембранного IgМ обусловлены особенностями считывания информации с генов локуса тяжелой цепи, лежащих за Сm-геном. В частности, за этим геном расположены короткие гены – Ск и Мк, кодирующие короткие аминокислотные последовательности дистальной части тяжелой цепи соответственно секреторного иммуноглобулина (ген Ск) и мембранного иммуноглобулина (ген Мк). При образовании секреторной формы IgМ, после считывания информации с Сm-гена транскрибируется только ген Ск, кодирующий концевой участок тяжелой цепи секреторной формы. При образовании тяжелых цепей мембранного IgМ транскрибируется более длинный участок генома за Сm-геном, включающий гены и Ск и Мк. Затем в результате процессинга участок мРНК, комплементарный Ск-гену, вырезается (подвергается делеции), а остальная часть длинной мРНК, кодирующей тяжелую цепь m-типа, путем сплайсинга сшивается с коротким фрагментом мРНК, комплементарным Мк-гену, в результате чего возникает зрелая мРНК, кодирующая мембранную форму IgМ. Таким образом, трансляция тяжелой цепи мембранного иммуноглобулина М представляет собой более сложный процесс, требующий больших энергозатрат, чем секреторной формы, поскольку предполагает считывание информации с более длинного участка генома и последующий процессинг и сплайсинг пре-мРНК, в связи с чем мембранной формы иммуноглобулина в В-лимфоцитах образуется гораздо меньше, чем секреторной. Синтез мембранной формы IgМ начинается на заключительных этапах дифференцировки В-лимфоцитов в красном костном мозге. Зрелые В-лимфоциты экспрессируют достаточно большое количество мембранного IgМ на своей поверхности: 2х105 молекул на клетку. Мембранная форма иммуноглобулина М на поверхности В-лимфоцитов может непосредственно взаимодействовать с любыми формами антигенов: как свободными корпускулярными (вирусами, бактериями) и молекулярными, так и связанными с рецепторами антигенпредставляющих клеток (моноцитов, макрофагов, дендритных клеток). Но в то же время молекулы мембранного IgМ сохраняют свою специфичность: молекулы каждого клона В-лимфоцитов проявляют сродство к строго определенным антигенным детерминантам, оставаясь равнодушными к другим.
Мембранная форма иммуноглобулина М отличается от секреторной не только тем, что является мономерной, но и некоторыми особенностями строения самого мономера IgМ: в его состав входит гидрофобный трансмембранный участок и короткий гидрофильный цитоплазматический хвост. В то же время мономерная молекула секреторной формы иммуноглобулина М имеет на С-конце углеводные остатки, увеличивающие растворимость молекулы, и цистеиновый остаток, необходимый для полимеризации мономеров в пентамерную форму с помощью J-цепи. Молекула мембранного иммуноглобулина М кодируется теми же рекомбинантными иммуноглобулиновыми локусами, что и секреторного иммуноглобулина (χ- или l-локусом, кодирующим легкую цепь, и локусом тяжелых цепей). Некоторые же отличия в строении С-конца тяжелых цепей секреторного и мембранного IgМ обусловлены особенностями считывания информации с генов локуса тяжелой цепи, лежащих за Сm-геном. В частности, за этим геном расположены короткие гены – Ск и Мк, кодирующие короткие аминокислотные последовательности дистальной части тяжелой цепи соответственно секреторного иммуноглобулина (ген Ск) и мембранного иммуноглобулина (ген Мк). При образовании секреторной формы IgМ, после считывания информации с Сm-гена транскрибируется только ген Ск, кодирующий концевой участок тяжелой цепи секреторной формы. При образовании тяжелых цепей мембранного IgМ транскрибируется более длинный участок генома за Сm-геном, включающий гены и Ск и Мк. Затем в результате процессинга участок мРНК, комплементарный Ск-гену, вырезается (подвергается делеции), а остальная часть длинной мРНК, кодирующей тяжелую цепь m-типа, путем сплайсинга сшивается с коротким фрагментом мРНК, комплементарным Мк-гену, в результате чего возникает зрелая мРНК, кодирующая мембранную форму IgМ. Таким образом, трансляция тяжелой цепи мембранного иммуноглобулина М представляет собой более сложный процесс, требующий больших энергозатрат, чем секреторной формы, поскольку предполагает считывание информации с более длинного участка генома и последующий процессинг и сплайсинг пре-мРНК, в связи с чем мембранной формы иммуноглобулина в В-лимфоцитах образуется гораздо меньше, чем секреторной. Синтез мембранной формы IgМ начинается на заключительных этапах дифференцировки В-лимфоцитов в красном костном мозге. Зрелые В-лимфоциты экспрессируют достаточно большое количество мембранного IgМ на своей поверхности: 2х105 молекул на клетку. Мембранная форма иммуноглобулина М на поверхности В-лимфоцитов может непосредственно взаимодействовать с любыми формами антигенов: как свободными корпускулярными (вирусами, бактериями) и молекулярными, так и связанными с рецепторами антигенпредставляющих клеток (моноцитов, макрофагов, дендритных клеток). Но в то же время молекулы мембранного IgМ сохраняют свою специфичность: молекулы каждого клона В-лимфоцитов проявляют сродство к строго определенным антигенным детерминантам, оставаясь равнодушными к другим.
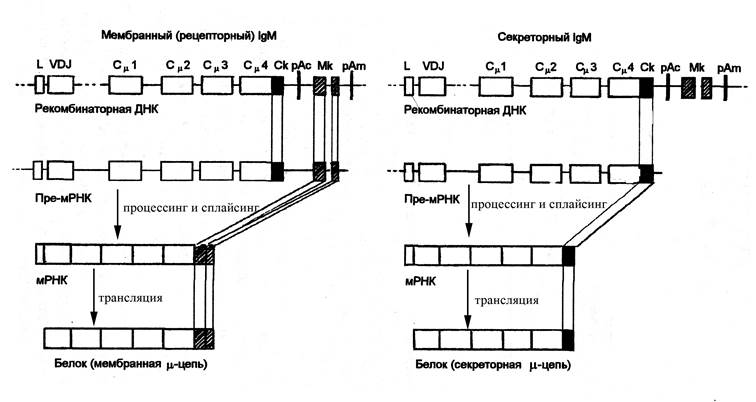
Рис. Схема генетического контроля синтеза мембранного и секреторного IgM. При образовании мембранной или секреторной форм IgM процесс реорганизации генома В-лимфоцита характеризуется следующими особенностями. Рекомбинантная ДНК содержит последовательность нуклеотидов от лидерного участка (L) до терминальной последовательности полиаденина (рАm – полиаденин мембранного пути развития) включительно. В реорганизованный локус ДНК входят экзон Ск (секреторный компонент) и два экзона Мк (мембранный компонент), один из которых контролирует трансмембранную последовательность, а второй – цитоплазматическую последовательность аминокислотных остатков рецепторного иммуноглобулина. В том случае, когда образующийся первичный транскрипт (пре-мРНК) включает экзоны Мк, синтезируется μ-цепь мембранного иммуноглобулина. При этом Ск и рАс (рАс – полиаденин секреторного пути реорганизации) подвергаются делеции при сплайсинге пре-мРНК. Если первичный транскрипт не включает последовательность Мк и рАm, но сохраняет Ск и рАс, то в результате последующих преобразований синтезируется μ-цепь секреторного иммуноглобулина
Взаимодействие мембранной формы IgМ В-лимфоцитов с определенным антигеном сопровождается активацией В-лимфоцитов, в результате которой В-лимфоциты превращаются в антителопродуцирующие клетки (плазматические клетки). Активация В-лимфоцита может происходить как самостоятельно после взаимодействия антигена с рецептором, так и зачастую с помощью цитокинов, продуцируемых активированными Т-хелперами. Вместе с тем сам по себе мембранный иммуноглобулин М после взаимодействия с антигеном активировать В-лимфоцит не может, поскольку имеет очень короткий цитоплазматический хвост, включающий всего несколько аминокислотных остатков, который не способен вступать в реакционные отношения с внутриклеточными ферментами и процессами. Для активации же В-лимфоцитов после присоединения к их антигенраспознающим рецепторам антигенов служат специальные вспомогательные низкомолекулярные белки – Iga и Igb, расположенные в непосредственной близости от мембранного IgМ в плазматической мембране В-лимфоцита и имеющие гораздо более длинные цитоплазматические концы. Эти белки после присоединения к мембранному IgМ антигенных эпитопов несколько изменяют свою конформацию, в результате чего становятся способными взаимодействовать с внутриклеточными тирозинкиназами, активируя их. Активированные тирозинкиназы, в свою очередь, активируют Са2+-зависимые фосфорилазы, катализирующие фосфорилирование определенных внутриклеточных белков. Образующиеся фосфорилированные белки влияют на конформацию связанных с ДНК белков таким образом, что усиливают транскрипцию специфических генов.
Большинство антигенов являются тимусзависимыми и для активации В-лимфоцитов требуют участия Т-хелперов. Тимуснезависимыми антигенами являются, как правило, полисахариды бактерий, имеющие множество повторяющихся одинаковых антигенных эпитопов, которые перекрестно сшиваются с большим количеством молекул мембранного IgМ на поверхности одного В-лимфоцита, что приводит к активации большого количества тирозинкиназ внутри В-лимфоцита и последующей его активации без участия Т-хелперов.
Дата добавления: 2016-07-18; просмотров: 3454;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
