Абиотические факторы
Свет
Свет является одним из основных абиотических факторов, влияющих на жизнедеятельность растений. Только на свету происходит фотосинтез и, следовательно, могут существовать фотосинтезирующие растения.
Следует помнить, что в растении непрерывно, вне зависимости от условий освещенности идет процесс дыхания, противоположный ассимиляции. При слабой интенсивности освещения количество энергии, затрачиваемое растением на дыхание, превышает то количество солнечной энергии, которое может быть связано растением в процессе фотосинтеза. Та интенсивность освещения, при которой расход энергии при дыхании полностью компенсируется энергией, запасаемой в процессе фотосинтеза, носит название компенсационной точки. У разных видов растений, в зависимости от их приспособленности к условиям окружающей среды, значение компенсационной точки может сильно различаться.
В зависимости от требовательности растений к количеству света, а также от влияния света на анатомо-морфологические особенности растительного организма выделяется 4 экологические группы растений.
Гелиофиты – светолюбивые растения, у которых компенсационная точка находится высоко, то есть процесс фотосинтеза начинает преобладать над процессом дыхания только при высокой интенсивности освещения и при этом не происходит его ингибирования при избыточном освещении. Для гелиофитов характерен замедленный рост побегов, низкая активность роста почек возобновления, в результате чего формируются низкорослые, компактные растения, а в горах – подушковидные. Листья, как правило, толстые, плотные, клетки эпидермиса не имеют хлоропластов, верхний эпидермис с толстой кутикулой, у горизонтально расположенных листьев мезофилл листа четко дифференцирован на столбчатую и губчатую паренхиму, межклетники не  большие, хлоропласты в клетках мелкие, многочисленные (рис. 5).
большие, хлоропласты в клетках мелкие, многочисленные (рис. 5).
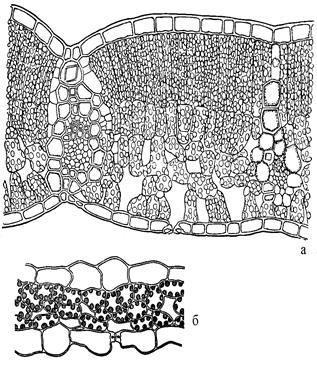 Сциофиты – тенелюбивые растения, у которых процесс фотосинтеза преобладает над процессом дыхания уже при слабой освещенности, тогда как высокая интенсивность солнечного света приводит к ингибированию фотосинтеза. Эти растения, как правило, могут довольствоваться короткими промежутками времени, когда интенсивность фотосинтеза возрастает благодаря падению на растение солнечного блика, тогда как все остальное время освещенность может быть ниже компенсационной точки (рис. 6).
Сциофиты – тенелюбивые растения, у которых процесс фотосинтеза преобладает над процессом дыхания уже при слабой освещенности, тогда как высокая интенсивность солнечного света приводит к ингибированию фотосинтеза. Эти растения, как правило, могут довольствоваться короткими промежутками времени, когда интенсивность фотосинтеза возрастает благодаря падению на растение солнечного блика, тогда как все остальное время освещенность может быть ниже компенсационной точки (рис. 6).
Для сциофитов характерны тонкие листья, клетки эпидермиса которых часто содержат хлоропласты, верхний эпидермис с тонкой кутикулой, мезофилл листа очень слабо или вообще не дифференцирован на столбчатую и губчатую паренхиму, межклетники крупные, хлоропласты в клетках крупные, их немного (рис. 5).
Умбропатиенты (факультативные сциофиты) – теневыносливые растения, которые находятся в оптимальных условиях при достаточно высокой интенсивности освещения, но, тем не менее, способные в течение длительного времени выносить сильное затенение. В большинстве своем это виды, обладающие высокой экологической 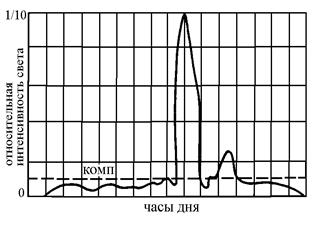 пластичностью. Фотосинтез у таких растений начинает преобладать над дыханием уже при низких значениях освещенности, но при увеличении интенсивности света не происходит ингибирования фотосинтеза, в отличие от типичных сциофитов.
пластичностью. Фотосинтез у таких растений начинает преобладать над дыханием уже при низких значениях освещенности, но при увеличении интенсивности света не происходит ингибирования фотосинтеза, в отличие от типичных сциофитов.
 Агелиофиты – растения, не нуждающиеся в свете (некоторые паразиты, сапротрофы).
Агелиофиты – растения, не нуждающиеся в свете (некоторые паразиты, сапротрофы).
 Помимо общего количества света, получаемого в течение дня, для растений также очень важно чередование дня и ночи в течение суток (фотопериодизм). Фотопериодизм закономерно меняется от экватора к умеренным и холодным зонам. Так, в районе экватора день практически равен ночи в течение всего года, тогда как в высоких широтах летом день значительно превышает ночь, а зимой – наоборот. Растения умеренных и холодных зон (растения длинного дня) в период своего роста нуждаются в высокой продолжительности светлого периода, для того, чтобы накопить достаточное количество органического вещества. Если их поместить в условия короткого дня, то они будут вегетировать, но не смогут перейти к цветению и плодоношению, так как будут испытывать постоянный недостаток в пластических веществах для образования цветков и плодов. Растения короткого дня, перенесенные в условия длинного светового дня, также не смогут перейти от вегетативного развития к генеративному. Это связано с тем, что в данном случае короткого темного периода будет недостаточно для оттока всех накопленных органических веществ из листьев и подготовки ассимиляционного аппарата к фотосинтезу. При этом продолжительность дня и ночи может быть одним из факторов, определяющих северную и южную границы распространения некоторых видов.
Помимо общего количества света, получаемого в течение дня, для растений также очень важно чередование дня и ночи в течение суток (фотопериодизм). Фотопериодизм закономерно меняется от экватора к умеренным и холодным зонам. Так, в районе экватора день практически равен ночи в течение всего года, тогда как в высоких широтах летом день значительно превышает ночь, а зимой – наоборот. Растения умеренных и холодных зон (растения длинного дня) в период своего роста нуждаются в высокой продолжительности светлого периода, для того, чтобы накопить достаточное количество органического вещества. Если их поместить в условия короткого дня, то они будут вегетировать, но не смогут перейти к цветению и плодоношению, так как будут испытывать постоянный недостаток в пластических веществах для образования цветков и плодов. Растения короткого дня, перенесенные в условия длинного светового дня, также не смогут перейти от вегетативного развития к генеративному. Это связано с тем, что в данном случае короткого темного периода будет недостаточно для оттока всех накопленных органических веществ из листьев и подготовки ассимиляционного аппарата к фотосинтезу. При этом продолжительность дня и ночи может быть одним из факторов, определяющих северную и южную границы распространения некоторых видов.
Частично отражая, частично поглощая и частично пропуская через себя, растение способно изменять интенсивность освещения и, что не менее важно, его спектральный состав. Таким образом, растения дифференцируют световые условия. К примеру, у древесных растений это зачастую приводит к тому, что часть ветвей в кроне не получает света в достаточном количестве и соответствующего качества, что в конечном итоге приводит к отмиранию нижних ветвей кроны (рис. 7). Количество света, проникающее сквозь крону деревьев, при этом будет тесно связано со значением компенсационной точки. Так, для лиственницы европейской (Larix decidua) относительный минимум силы света составляет примерно 20% от полного дневного освещения, а для ели европейской (Picea abies) – лишь 3%. Именно эти значения и будут определять световой режим под пологом данных видов деревьев.
Тепло
Тепло является одним из наиболее важных экологических факторов, который определяет распределение растений по земному шару. Общий температурный диапазон активной жизни для растений на поверхности нашей планеты колеблется в довольно узких рамках – от +55 ºС до –70 ºС. Причем, помимо общего количества тепла, очень важное значение имеет также распределение его во времени, т. е. температурный режим. На суше температурный режим сравнительно постоянен лишь в тропиках, на остальной же территории планеты он изменяется в двух направлениях: от тропиков к полярным широтам уменьшается общее количество тепла, поступающего на поверхность Земли, а при продвижении от побережья вглубь континентов возрастают амплитуды колебаний суточных и годовых температур.
Все растения по отношению к температуре окружающей среды можно разделить на две группы: эвритермные и стенотермные. Эвритермные растения имеют широкую экологическую амплитуду по отношению к этому фактору и, следовательно, способны существовать при больших колебаниях температуры среды. Способствуют этому, как правило, различные физиологические механизмы адаптации. Это выражается в особенностях химического состава протоплазмы (в частности, в ее способности изменять концентрацию растворимых веществ в широких пределах), теплоустойчивости ферментов, положении оптимума их активности и др. Классическим примером эвритермного вида может быть мятлик однолетний (Poa annua) – вид-космополит, широко распространенный от тундровой зоны и до субтропиков. Стенотермные растения приспособлены к относительно постоянным температурным условиям и не переносят их сильного колебания. Стенотермными являются многие тропические растения. Так банан (Musa x paradisiaca) при понижении температуры ниже +16 ºС практически прекращает свой рост.
Растения и, тем более, растительные сообщества зачастую способны оказывать существенное влияние на суточный, сезонный и годовой ход температур. Особенно сильно подобное влияние выражено в лесных сообществах. Так, под пологом леса уменьшаются суточные и сезонные амплитуды температур, то есть температурный фон становится более ровным. По сравнению с открытыми пространствами в лесах несколько ниже средние суточные и годовые температуры, что связано, в первую очередь, с перехватом и отражением значительной части солнечной энергии кронами деревьев.
Вода
Вода как экологический фактор для растений может иметь значение в виде атмосферных осадков (их количества, формы (дождь, туман, роса, снег и др.), продолжительности, распределения во времени), а также в виде содержания водяного пара в атмосфере. Не следует забывать и о том, что лимитирующее воздействие на растения фактора влажности может в очень сильной степени зависеть от воздействия других абиотических факторов, особенно если они имеют критические значения. К примеру, низкая температура может препятствовать всасыванию воды из почвы, тогда как излишне высокая – приводить к чрезмерной ее транспирации растениями.
По отношению к воде как к экологическому фактору выделяются следующие группы растений:
· ксерофиты – растения сухих местообитаний, которые могут переносить перегрев и обезвоживание. Ксерофиты в зависимости от способа перенесения неблагоприятных условий подразделяются не несколько подгрупп:
o эуксерофиты хорошо переносят длительное обезвоживание и перегрев, так как их протоплазма имеет очень высокое осмотическое давление, а обмен веществ малоинтенсивен. Они имеют сравнительно неглубоко расположенную корневую систему и ксероморфное строение надземных органов (опушение, плотные сильно кутинизированные покровные ткани, в различной степени редуцированные листья и др.);
o гемиксерофиты устойчивы к засухе благодаря мощной корневой системе, достигающей грунтовых вод, интенсивным процессам транспирации и обмена веществ; не выносят обезвоживания;
o суккуленты устойчивы к засухе благодаря сильному развитию водозапасающей паренхимы в стебле, листьях или корне, клетки которой имеют большую вязкость протоплазмы и содержат большое количество связанной воды, а также плотным, часто кутинизированным покровным тканям; не выносят обезвоживания;
o пойкилоксерофиты при обезвоживании впадают в анабиоз;
o криофиты – растения, приспособленные к сухим холодным местообитаниям, например, высокогорьям;
o психрофиты – ксероморфные растения, произрастающие на влажных холодных почвах; дефицит влаги у них связан с низкими температурами;
· мезофиты – растения, обитающие в условиях с более или менее достаточным, но не избыточным количеством воды в почве. Среди мезофитов выделяют особую группу эфемеров (однолетние растения) и эфемероидов (многолетники), вегетация которых длится всего несколько недель и они, зачастую обитая в засушливых зонах, для вегетации используют период с достаточной водообеспеченностью;
· гигрофиты – сухопутные растения, произрастающие в условиях высокой влажности грунта и воздуха; у них отсутствуют приспособления, ограничивающие расходование воды и слабо развитая корневая система, поэтому даже незначительный недостаток воды вызывает их увядание;
· гидрофиты – водные растения, прикрепленные к грунту и погруженные в воду только нижними частями, с хорошо развитыми корнями, механическими тканями и сосудами.
· гидатофиты – водные растения, целиком или большей частью погруженные в воду, с очень слабо развитыми или, иногда, отсутствующими корневой системой, механическими тканями и сосудами.
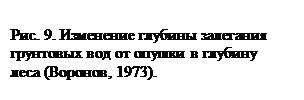

 При анализе распределения осадков на поверхности нашей планеты выявляются определенные закономерности. Так, наибольшее количество осадков выпадает на экваторе, по направлению к тропикам и субтропикам их количество падает, затем вновь возрастает в умеренных зонах и вновь падает в Арктике и Антарктике. Это зависит от характера циркуляции атмосферы на нашей планете: устойчивый восходящий ток теплого и влажного воздуха в районе экватора, нисходящий ток жаркого сухого воздуха в тропиках, а также ход циклонов в умеренных зонах. Помимо характера циркуляции атмосферы, значительную роль в определении количества годовых осадков играет расстояние от теплого моря: чем оно ближе, тем больше осадков выпадает. Немаловажную роль в перераспределении осадков играют горные системы, которые способны препятствовать нормальной циркуляции атмосферы над сушей. Результатом этого часто бывает чрезмерное количество осадков с одной стороны горной системы и недостаток – с другой.
При анализе распределения осадков на поверхности нашей планеты выявляются определенные закономерности. Так, наибольшее количество осадков выпадает на экваторе, по направлению к тропикам и субтропикам их количество падает, затем вновь возрастает в умеренных зонах и вновь падает в Арктике и Антарктике. Это зависит от характера циркуляции атмосферы на нашей планете: устойчивый восходящий ток теплого и влажного воздуха в районе экватора, нисходящий ток жаркого сухого воздуха в тропиках, а также ход циклонов в умеренных зонах. Помимо характера циркуляции атмосферы, значительную роль в определении количества годовых осадков играет расстояние от теплого моря: чем оно ближе, тем больше осадков выпадает. Немаловажную роль в перераспределении осадков играют горные системы, которые способны препятствовать нормальной циркуляции атмосферы над сушей. Результатом этого часто бывает чрезмерное количество осадков с одной стороны горной системы и недостаток – с другой.
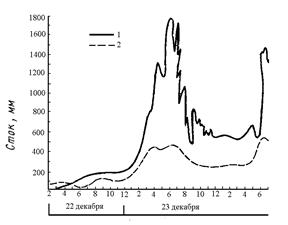
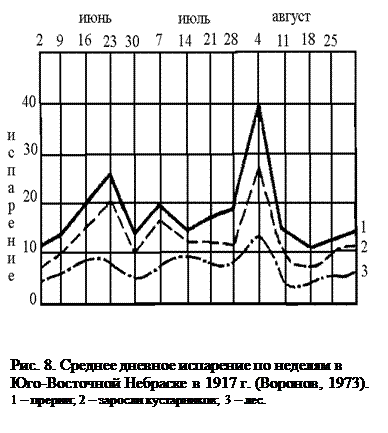
Осадки, выпадающие над сушей, могут образовывать поверхностный и внутрипочвенный сток, испаряться, а также транспирироваться растениями. На все эти процессы растения оказывают существенное влияние. Так, в различных типах растительности с поверхности почвы испаряется различное количество влаги (рис. 8). Как видно из приведенного графика, в лесных сообществах по сравнению с открытыми участками испарение с поверхности почвы значительно уменьшается, чему в значительной степени способствует лесная подстилка. Однако если сравнивать общее количество влаги, возвращающееся в атмосферу, то закономерность будет прямо противоположной. Дело в том, что в лесу значительная доля осадков вообще не достигает поверхности почвы, задерживаясь кронами деревьев. Вода, испаряясь с поверхности листьев, ветвей и стволов, увеличивает относительную влажность воздуха в лесу по сравнению с открытыми пространствами. Именно поэтому в лесных сообществах до 1/3 влаги, выпадающей над ними в виде осадков, может возвращаться в атмосферу, вообще не попадая в почву. Кроме того, деревья, имея, по сравнению с травянистыми растениями, большую листовую поверхность, транспирируют значительно большее количество воды, понижая при этом уровень грунтовых вод (рис. 9). Лесные сообщества замедляют также поверхностный сток воды во время сильных дождей (рис. 10). Все это приводит к тому, что над облесенными территориями суши выпадает большее количество осадков, чем над открытыми.
Воздух
Для растений наиболее важны такие характеристики воздушного режима, как температура, влажность, химический состав и скорость движения воздушных масс. Первые 2 характеристики нами уже кратко рассматривались выше.
Химический состав воздуха. Наибольшее влияние на растения оказывают содержание в воздухе углекислого газа и различных примесей. Загрязнение воздуха может быть очень важным для растений в районах с высокоразвитой индустрией. Так, крайне негативное влияние на рост и развитие растений оказывает содержание в воздухе соединений серы (чаще всего оксидов), причем подавление роста растений происходит уже при концентрации диоксида серы, равной 0,0002-0,0005 %. Отрицательное влияние может оказывать и степень запыленности атмосферы, так как пыль, покрывающая листья растений, может препятствовать нормальному протеканию фотосинтеза. Повышенная концентрация в воздухе некоторых соединений азота (например, аммиака), наоборот, может стимулировать рост растений, но только в том случае, если она не слишком большая и не превышает порога токсичности.
 Движение воздуха (ветер). Влияние ветра на жизнь и развитие растений сложно и многообразно. Ветер является важным фактором распространения: он переносит споры, семена, плоды растений. В конечном итоге именно способность тех или иных видов растений к распространению в значительной мере определяет флористический состав растительных сообществ, формирующихся на нарушенных территориях. Очень важна роль ветра в процессах опыления, так как именно с помощью ветра переносится пыльца всех голосеменных и многих покрытосеменных растений. Ветер приводит к тому, что вокруг растения постоянно происходит смена воздуха. С одной стороны, это полезно для растения, так как к нему приносятся новые дозы углекислого газа. С другой стороны, ветер действует на растения иссушающе, так как воздух вокруг растения, обогащенный выделенным им водяным паром, заменяется более сухим. Ветер охлаждает растение, однако, если воздух несет большое количество сильно нагретых твердых частиц (пыль, песок), то в таком случае он может, наоборот, приводить к перегреву растения. Постоянное однонаправленное действие ветра значительной силы может приводить к значительному изменению внешнего облика растений. Особенно ярко это проявляется у одиночно стоящих деревьев, произрастающих на берегу моря или в горах, где преобладают ветры одного направления и зачастую значительной силы. В таком случае иссушающее действие ветра будет значительно сильнее проявляться с наветренной стороны кроны, где происходит замедление роста ветвей или они вовсе не развиваются. При этом у деревьев формируется односторонняя, асимметричная крона (рис. 11). Сильный ветер способен обламывать отдельные ветви деревьев и даже выворачивать их с корнем из земли. Это приводит к изменению светового режима внутри растительного сообщества и формированию новых экологических ниш.
Движение воздуха (ветер). Влияние ветра на жизнь и развитие растений сложно и многообразно. Ветер является важным фактором распространения: он переносит споры, семена, плоды растений. В конечном итоге именно способность тех или иных видов растений к распространению в значительной мере определяет флористический состав растительных сообществ, формирующихся на нарушенных территориях. Очень важна роль ветра в процессах опыления, так как именно с помощью ветра переносится пыльца всех голосеменных и многих покрытосеменных растений. Ветер приводит к тому, что вокруг растения постоянно происходит смена воздуха. С одной стороны, это полезно для растения, так как к нему приносятся новые дозы углекислого газа. С другой стороны, ветер действует на растения иссушающе, так как воздух вокруг растения, обогащенный выделенным им водяным паром, заменяется более сухим. Ветер охлаждает растение, однако, если воздух несет большое количество сильно нагретых твердых частиц (пыль, песок), то в таком случае он может, наоборот, приводить к перегреву растения. Постоянное однонаправленное действие ветра значительной силы может приводить к значительному изменению внешнего облика растений. Особенно ярко это проявляется у одиночно стоящих деревьев, произрастающих на берегу моря или в горах, где преобладают ветры одного направления и зачастую значительной силы. В таком случае иссушающее действие ветра будет значительно сильнее проявляться с наветренной стороны кроны, где происходит замедление роста ветвей или они вовсе не развиваются. При этом у деревьев формируется односторонняя, асимметричная крона (рис. 11). Сильный ветер способен обламывать отдельные ветви деревьев и даже выворачивать их с корнем из земли. Это приводит к изменению светового режима внутри растительного сообщества и формированию новых экологических ниш.
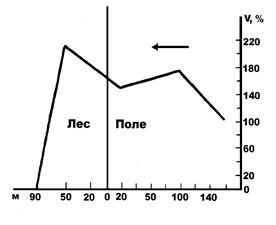
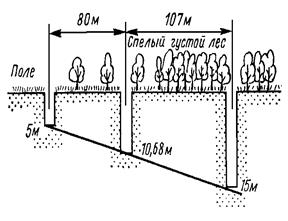
 Растения в свою очередь также существенно изменяют движение ветра, действуя как механическое препятствие при перемещении воздушных масс. Это в первую очередь сказывается на изменении скорости ветра. К примеру, в лесу уже на расстоянии 90 м от опушки скорость ветра составляет лишь несколько процентов от первоначальной (рис. 12), хотя стоит отметить, что это расстояние может очень сильно варьировать в зависимости от строения опушки: чем она плотнее, тем быстрее замедляется скорость ветра. Действие трав и кустарничков на ветер в принципе имеет тот же характер, но они гасят ветер в припочвенном слое. Так, заросли вереска способны замедлить ветер почти в десять раз: если над вершинами кустиков вереска скорость ветра составляет 9,3 м/с, то в куртинах на высоте 50 см падает до 3,7 м/с, а на высоте 10 см над поверхностью почвы – только 1 м/с (Ипатов, Кирикова, 1999).
Растения в свою очередь также существенно изменяют движение ветра, действуя как механическое препятствие при перемещении воздушных масс. Это в первую очередь сказывается на изменении скорости ветра. К примеру, в лесу уже на расстоянии 90 м от опушки скорость ветра составляет лишь несколько процентов от первоначальной (рис. 12), хотя стоит отметить, что это расстояние может очень сильно варьировать в зависимости от строения опушки: чем она плотнее, тем быстрее замедляется скорость ветра. Действие трав и кустарничков на ветер в принципе имеет тот же характер, но они гасят ветер в припочвенном слое. Так, заросли вереска способны замедлить ветер почти в десять раз: если над вершинами кустиков вереска скорость ветра составляет 9,3 м/с, то в куртинах на высоте 50 см падает до 3,7 м/с, а на высоте 10 см над поверхностью почвы – только 1 м/с (Ипатов, Кирикова, 1999).
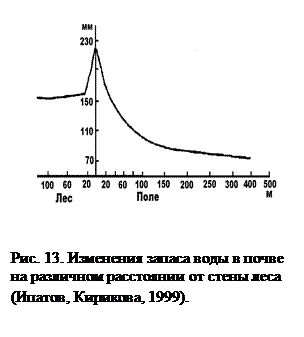
Растения, замедляя движение воздуха, препятствуют также переносу ветром снега. Это приводит к тому, что снег, сдуваемый с открытых пространств, скапливается в приопушечной зоне. Таким образом, весной запасы влаги на опушке леса будут значительно больше, чем на открытом пространстве или же в глубине лесного массива (рис. 13).
Почвы и грунты
Эдафические условия определяются свойствами субстрата, на котором произрастают растения. Субстрат может быть лишь незначительно затронут процессами почвообразования (скалы, подвижные пески), или же в результате действия физического выветривания и живых организмов поверхностный слой горной породы может превратиться в почву.
Почвы играют для растений крайне важное значение. Во-первых, в большинстве случаев почвы являются тем субстратом, в котором происходит механическое закрепление растений. Во-вторых, именно почвы обеспечивают водоснабжение и минеральное питание растений. В третьих, через почвы осуществляются многие типы взаимодействий растений друг с другом, а также с другими организмами (почвенными микроорганизмами, грибами, животными). Почвы характеризуются рядом физико-химических характеристик, имеющих существенное значение для роста и развития растений, а также для формирования того или иного типа растительного сообщества.
Воздушный режим почв определяет обеспеченность корней растений кислородом, необходимым для дыхания. Он зависит от механического состава почв: чем большие размеры имеют частицы субстрата, тем большая воздухопроницаемость. К примеру, высокая воздухопроницаемость характерна для песчаных грунтов, а для глинистых – значительно более низкая.
Водный режим грунтов определяет запасы доступной для растения почвенной влаги. Всю воду, находящуюся в почве, можно разделить на две категории: доступная для растений капиллярная влага и недоступная физически и химически связанная влага. Но даже если в почве имеется влага в достаточном или избыточном количестве, она не всегда может быть доступна для растений. В этом случае говорят о так называемой физиологической сухости почвы. Вызывать это явление могут несколько абиотических факторов. Так, физиологически сухими являются богатые влагой, но имеющие очень низкую температуру почвы тундр. На верховых болотах с обильным сфагновым покровом вода оказывается недоступной для многих видов из-за ее низкой температуры, высокой кислотности, слабой аэрации и наличия токсических веществ. Все это в совокупности нарушает функционирование корневой системы растений. Физиологически сухими являются также и засоленные почвы, в которых влага благодаря высокому осмотическому давлению почвенного раствора не может быть использована растениями. Физиологически сухие местообитания экстремальны, и поэтому произрастающие в них растения должны быть приспособлены либо к повышенному содержанию в почве легкорастворимых солей, либо к очень низкой температуре почвы.
Кислотность почвы, так же как и ее физические особенности, сильно влияет на рост и развитие растений. Кислотность почвы выражается отрицательным логарифмом концентрации водородных ионов рН. При рН почвы в диапазоне 2,4-6,7 она считается кислой, 6,7-7,0 – нейтральной, выше 7,0 – щелочной. Большинство почв Беларуси являются кислыми.
Кислотность грунтов может быть вызвана многими причинами: химическим составом материнских пород, выделений растений и растительных остатков, степенью аэрации и др. Так, кислые грунты формируются в условиях чрезмерного увлажнения. На гранитах, гнейсах, андезитах также формируются кислые почвы, тогда как на лёссах, мелах, мергелях – нейтральные или щелочные. Почвы под еловыми лесами, по сравнению с широколиственными, будут иметь более кислую реакцию, так как продукты разложения растительных остатков хвойных пород (хвоя, древесина) имеют более кислую реакцию.
По отношению к кислотности почвы как к экологическому фактору выделяются следующие группы растений (Braun-Blanquet, 1951):
· ацидофилы – растения, предпочитающие кислые почвы (рН 2,4-6,7). В пределах этой группы выделяются экологические подгруппы с более узкими значениями, которые точнее характеризуют природные условия: крайние ацидофилы (рН 2,4-5,0), умеренные ацидофилы (рН 5,0-6,2) и слабые ацидофилы (рН 6,2-6,7);
· нейтрофилы – растения нейтральных почв (рН 6,7-7,0). Это чина луговая (Lathyrus pratensis), тимофеевка луговая (Phleum pratense), ежа сборная (Dactylis glomerata) и др.;
· базифилы – растения щелочных почв (рН 7,0 и выше). Их можно разделить на слабые базифилы (рН 7,0-8,0) и выраженные базифилы (рН 8,0-14,0, как правило, это растения, произрастающие на мелах и имеющие довольно узкое распространение);
· индифферентные виды не имеют определенной приуроченности к тому или иному значению рН почвы и встречаются в очень широких пределах рН – начиная от щелочных и кончая сильнокислыми. Типичными представителями этой группы являются ландыш майский (Convallaria majalis), сосна обыкновенная (Pinus sylvestris), дуб черешчатый (Quercus robur) и др.
Валовое содержание питательных веществ (богатство почвы) определяет плодородие почвы и, следовательно, уровень обеспечения растений питательными веществами. Отсутствие или недостаток того или иного элемента питания (азота, калия, фосфора, кальция, железа, магния, других макро- и микроэлементов) приводит к нарушениям метаболизма растений. К примеру, при недостатке магния, являющегося структурным компонентом хлорофилла, последний разрушается, что приводит к нарушениям процессов фотосинтеза.
По отношению к богатству почвы как к экологическому фактору выделяются следующие группы растений:
· эвтрофы – растения, нормально развивающиеся только на почвах, богатых питательными веществами. Индикаторами богатства грунта могут служить крапива двудомная (Urtica dioica), чистотел большой (Chelidonium majus), липа (Tilia cordata), ясень (Fraxinus excelsior) и т.д. К этой группе относятся также нитрофилы – растения, нуждающиеся в повышенном содержании в почве азота. Они поселяются на свалках у жилья человека, около животноводческих ферм, на перегное в широколиственных лесах и др. Типичными нитрофилами являются многие виды лебеды (Atriplex), мари (Chenopodium), бузина красная (Sambucus racemosa), хмель (Humulus lupulus) и др.;
· мезотрофы – растения, обитающие на почвах с умеренным содержанием элементов минерального питания. К мезотрофам относятся многие культурные растения: морковь (Daucus sativus), яблоня (Malus domestica), картофель (Solanum tuberosum) и др., а также большая часть дикорастущих видов растений;
· олиготрофы – растения, мало требовательные к плодородию почвы и поэтому могущие произрастать на грунтах с минимальным содержанием доступного азота и зольных элементов. К олиготрофам относятся растения экстремальных местообитаний – верховых болот, песков, скал. Во флоре Беларуси типичными олиготрофами являются многие виды сфагновых мхов, овсяниц, булавоносец седой (Corynephorus canescens), вереск (Calluna vulgaris), вейник наземный (Calamagrostis epigeios), сосна (Pinus sylvestris) и др.
Содержание солей в почве. Процессы засоления почв могут идти в различных природно-климатических условиях при преобладании в почве восходящего тока воды, несущего с собой легкорастворимые соли, над нисходящим током. Благодаря этому перемещение солей из верхних горизонтов в нижние становится невозможным, и легкорастворимые соли постепенно накапливаются в верхних горизонтах грунта. Эти процессы характерны для сухих областей земного шара, где испарение резко превышает количество выпадающих осадков, а также по берегам океанов и морей.
Растения, приспособленные к высокому содержанию солей в почве, носят название галофитов. Они характеризуются высоким осмотическим давлением клеточного сока (у некоторых видов до 100 атм. и выше), что дает им возможность извлекать воду из концентрированного солевого раствора почвы. Большинство галофитов либо имеет суккулентное, либо ксероморфное строение. Растения незасоленных почв называются гликофитами.
Среди легкорастворимых солей, участвующих в засолении грунта в аридных областях, наиболее обычны соли натрия (NaCl, Na2SO4, NaHCO3 и Na2CO3), хлориды и сульфаты кальция и магния. Значительно реже встречаются соли калия и азотнокислые соли. На морских и океанических побережьях основным фактором засоления является хлорид натрия, количество которого в различных географических зонах может варьировать в широких пределах. Так, к примеру, на берегу Балтийского моря почвы могут иметь концентрацию хлористого натрия не выше, чем в питательном растворе Кноппа, на побережье Северного моря – до 6%, а у берегов океана в экваториальной зоне при интенсивном испарении воды во время приливов – до 12%.
Засоленные почвы делятся на два типа: солончаки и солонцы. Солончаки характеризуются высоким содержанием легкорастворимых солей у самой поверхности. На солончаках произрастают преимущественно типичные галофиты. В отличие от солончаков, солонцы содержат легкорастворимые соли на некоторой глубине (20-50 см и глубже), верхние же горизонты их промыты атмосферными осадками и почти лишены легкорастворимых солей. На солончаках произрастают главным образом растения-ксерофилы, приспособленные к существованию на сухих почвах. Типичные же галофиты на солонцах встречаются довольно редко.
От механического состава субстрата, на котором растут растения, в значительной степени зависят его воздушный, водный, температурный режимы и др. Так, песчаные почвы воздухо- и водопроницаемы, легко прогреваются и так же легко остывают. Это приводит к тому, что органика в таких почвах быстро минерализуется, и питательные вещества вместе с нисходящим током воды вымываются в нижние горизонты почвы. Глинистые почвы, наоборот, обладают сильной водоудерживающей способностью, их температурный режим более стабилен, восходящий и нисходящий ток более или менее уравновешен и основная масса питательных веществ сохраняется в верхних слоях почвы. Как мы видим, механически состав субстрата играет для растений чаще всего опосредованную роль. Однако существуют субстраты, где механический состав является основным лимитирующим фактором для растений: открытые пески и скалы.
Псаммофиты (растения подвижных песков) обладают целым рядом приспособлений, обеспечивающих им возможность произрастания на подвижных субстратах, имеющих рыхлую структуру. Они способны образовывать придаточные корни на засыпанных песком стеблях и придаточные почки на корнях, оказавшихся на поверхности в связи с выдуванием песка. У некоторых кустарников побеги способны очень быстро расти и ускользать от заноса песком. Многие виды имеют длинные быстрорастущие корневища, что позволяет им быстро расселяться и закреплять песок. Корни псаммофитов имеют многочисленные корневые волоски и одеты чехликами из частиц песка, надежно защищающими их от высыхания. Листья псаммофитов узкие, жесткие, а у многих видов полностью отсутствуют, что облегчает сопротивление ветру, нередко несущему тучи песка. Тем не менее, многие псаммофиты очень сильно транспирируют, что позволяет им уменьшать нагрев зеленых фотосинтезирующих частей. Воду при этом они получают за счет сильно развитой корневой системы. Плоды и семена псаммофитов имеют различные придатки, которые обеспечивают их распространение, быстрое заглубление в грунт, прорастание и укоренение молодых проростков.
Литофиты (петрофиты) – растения, приспособленные к жизни на камнях, скалах или в их трещинах. Обычно на камне сначала поселяются бактерии и водоросли, затем накипные лишайники, потом листоватые лишайники и мхи, и, наконец, высшие растения. Литофиты могут поселяться на поверхности камня, не разрушая его (эпилиты), способны разрушать горные породы химическими выделениями (особенно кислотами) своих ризоидов или корней (литофагофиты), или же поселяться на детрите и первичной почве в углублениях и трещинах скал (хазмофиты). В любом случае эти растения имеют ряд общих приспособлений, которые им позволяют расти в подобных экстремальных условиях. Так, подавляющее большинство литофитов – суккуленты (камнеломки), эуксерофиты (эдельвейсы, овсяницы, колокольчики и др.) или пойкилоксерофиты (большинство видов лишайников и мхов), зачастую способные впитывать воду всей поверхностью своего тела. Практически все литофиты являются олиготрофами, причем многие растения способны получать минеральные вещества прямо из камня, растворяя материнскую породу выделениями своих корневых волосков или ризоидов. Литофиты играют в природе очень важную роль, так как они являются пионерами, способствующими разрушению горных пород, их отмершие остатки создают основу для формирования гумуса и образования полноценной почвы.
Все вышеперечисленное очень важно для понимания взаимоотношений между растениями и почвенной средой. Выше кратко рассмотрено, как почвы влияют на рост, развитие растений и их распределение на поверхности планеты. Но не следует забывать и о том, что растения тоже влияют на почвы. Растительные остатки, перегнивая, формируют основу гумуса – бесструктурную органическую составляющую почвы. Возникновению структуры почвы во многом способствуют корни растений. Пронизывая толщу субстрата, они оказывают механическое давление и уплотняют его, то есть, сеть корней в определенной мере способствует обособлению структурных частиц почвы. Постоянное потребление воды растениями приводит к изменению водного режима почвы, дыхание корней растений влияет на состав почвенного воздуха и, следовательно, на состав почвенного раствора. Минеральное питание растений является обменным процессом: избирательно поглощая из почвы те или иные минеральные вещества в виде ионов, растение выделяет в почву эквивалентное количество других ионов (чаще всего Н+ и НСО3-). Кроме того, корни растений выделяют в почву большое количество органических веществ. Все это приводит к тому, что растения в процессе жизнедеятельности в той или иной степени меняют химический состав почвы. Поглощенные растениями минеральные вещества вместе с опадом (отмершие ветви, листья, плоды и др.) попадают на поверхность почвы или в самые верхние ее слои в составе сложных органических соединений, где они разрушаются сапротрофными организмами. Это приводит к перераспределению минеральных веществ в почве, то есть обогащению верхних слоев за счет более глубоких.
Как мы видим, почва с поселением на ней растений изменяется при активном воздействии последних. Причем различные виды растений и, следовательно, различные растительные сообщества в различной степени изменяют почвы. Это приводит к тому, что под разными растительными сообществами формируются почвы, различающиеся своим составом и структурой. Так, в условиях Беларуси в лесных сообществах без избыточного увлажнения формируются, как правило, подзолистые почвы. В то же время в аналогичных условиях на лугах формируются дерново-подзолистые почвы.
Рельеф
Орографические факторы (характер рельефа, высота над уровнем моря, протяженность, характер склонов и др.) – очень сложно, косвенно действующие факторы. Они не являются экологическими факторами в узком смысле, так как непосредственно на растительные сообщества влияния не оказывают. Тем не менее, роль рельефа как фактора, очень сильно влияющего на климатические и эдафические факторы, чрезвычайно велика. С рельефом связана вертикальная поясность растительного покрова. В большинстве случаев именно разнообразие условий рельефа обеспечивает разнообразие растительных сообществ в пределах одной климатической зоны. Нередко рельеф определяет распространение тех или иных видов растений и, тем самым, видовой состав растительных сообществ.
Принято различать четыре категории рельефа – макрорельеф, мезорельеф, микрорельеф и нанорельеф.
Макрорельеф – это крупные формы рельефа (горные хребты, плоскогорья, равнины, низины), образованные эндогенными процессами и определяющие основные особенности растительности крупного региона. Элементы макрорельефа создают на относительно небольшой площади настолько большую амплитуду высот, что возникает целая серия климатических комплексов. Это связано, в первую очередь с тем, что при подъеме на каждые 100 м температура понижается в среднем на 0,5-0,6 °С. Одновременно с изменением температуры меняются также режим осадков и условия освещения. Все это приводит к тому, что при подъеме в горы можно наблюдать такое явление, как вертикальная поясность. Вертикальная поясность часто схожа с изменениями растительности на равнине, которые можно наблюдать по мере движения от подножия данной горной системы в сторону полюса того же полушария (широтная зональность). Однако эта закономерность не абсолютна, так как в горах можно довольно часто наблюдать выпадение того или иного пояса растительности, инверсию поясов и др. Кроме того, распределение растительности на склонах разной экспозиции даже на одной и той же горе будет очень сильно отличаться.
Помимо формирования вертикальной поясности, макрорельеф также оказывает очень сильное влияние на растительность, преграждая путь ветрам, обладающим определенными свойствами (например, сухим или богатым влагой, теплым или холодным). Такое влияние может распространяться очень далеко за пределы расположения горной системы. Так, преграждающие путь влажным западным ветрам Кордильеры являются причиной того, что на североамериканском континенте природные зоны, начиная с ш<
Дата добавления: 2020-08-31; просмотров: 1870;
Поиск по сайту
Узнать еще

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
