Возникновение хромосомных аберраций
Основной предпосылкой для возникновения хромосомных перестроек является появление в клетке двунитевых разрывов ДНК, то есть разрывов обоих нитей спирали ДНК в пределах нескольких п.о. Двунитевые разрывы ДНК возникают в клетке спонтанно или под действием различных мутагенных факторов: физической (ионизирующее излучение), химической или биологической (транспозоны, вирусы) природы. Нарушения и ошибки процесса репарации двунитевых разрывов ДНК приводят к появлению хромосомных перестроек.
Классификация
Делеции
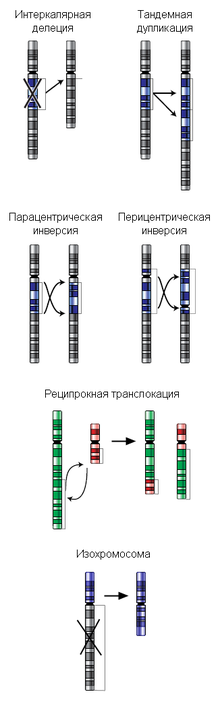
Некоторые типы хромосомных перестроек
Различают терминальные (утрата концевого участка хромосомы) и интеркалярные (утрата участка на внутреннем участке хромосомы) делеции. Если после образования делеции хромосома сохранила центромеру, она аналогично другим хромосомам передается при митозе, участки же без центромеры, как правило, утрачиваются. При конъюгации гомологичных хромосом во время мейоза у нормальной хромосомы на месте, соответствующем интеркалярной делеции у дефектной хромосомы, образуется делеционная петля, которая компенсирует отсутствие делетированного участка.
Врождённые делеции у человека редко захватывает протяженные участки хромосом, обычно такие аберрации приводят к гибели эмбриона на ранних этапах развития. Самым хорошоизученным заболеванием, обусловленным достаточно крупной делецией, является синдром кошачьего крика, описанный в 1963 годуЖеромом Леженом. В его основе лежит делеция участка короткого плеча 5 хромосомы. Для больных характерен ряд отклонений от нормы: нарушение функцийсердечно-сосудистой, пищеварительной систем, недоразвитие гортани (с характерным криком, напоминающим кошачье мяуканье), общее отставание развития,умственная отсталость, лунообразное лицо с широко расставленными глазами. Синдром встречается у 1 новорожденного из 50000.
Современные методы выявления хромосомных нарушений, прежде всего флуоресцентная гибридизация in situ, позволили установить связь между микроделециями хромосом и рядом врождённых синдромов. Микроделециями, в частности, обусловлены давно описанные синдром Прадера-Вилли исиндром Вильямса.
Дупликации
Дупликации представляют собой класс перестроек, который объединяет как внутри- , так и межхромосомные перестройки. Вообще, любая дупликация — это появление дополнительной копии участка хромосомы, которая может располагаться сразу за тем районом, который дуплицирован, тогда это тандемная дупликация, либо в новом месте или в другой хромосоме. Новая копия может образовать отдельную маленькую хромосому со своими собственными теломерами и центромерой, тогда это свободная дупликация[1]:2. Тандемные дупликации появляются в половых клетках при мейозе в результате неравного кроссинговера (в этом случае второй гомолог несет делецию) или в соматических клетках в результате неаллельной гомологичной рекомбинации при репарации двунитевого разрыва ДНК. В процессе кроссинговера у гетерозиготы при конъюгации хромосомы с тандемной дупликацией и нормальной хромосомы, как и при делеции, формируется компенсационная петля.
Практически у всех организмов в норме наблюдается множественность генов, кодирующих рРНК (рибосомальную РНК). Это явление назвали избыточностью генов. Так у E. coli на рДНК (ДНК, кодирующее рРНК) приходится 0,4 % всего генома, что соответствует 5-10 копиям рибосомальных генов.
Другой пример дупликации — мутация Bar у Drosophila, обнаруженная в 20-х годах XX века Т. Морганом и А. Стёртевантом. Мутация обусловлена дупликацией локуса 57.0 X-хромосомы. У нормальных самок (B+/B+) глаз имеет 800 фасеток, у гетерозиготных самок (B+/B) глаз имеет 350 фасеток, у гомозиготпо мутации (B/B) — всего 70 фасеток. Обнаружены также самки с трижды повторенным геном — double Bar (BD/B+).
В 1970 году Сусумо Оно в монографии «Эволюция путем дупликации генов» разработал гипотезу об эволюционной роли дупликаций, поставляющих новые гены, не затрагивая при этом функций исходных генов. В пользу этой идеи говорит близость ряда генов по нуклеотидному составу, кодирующих разные продукты. Это трипсин и химотрипсин, гемоглобин и миоглобин и ряд других белков.
Инверсии
Инверсией называют поворот участка хромосомы на 180°. Различают парацентрические (инвертированный фрагмент лежит по одну сторону от центромеры) и перицентрические (инвертированный фрагмент лежит по разные стороны от центромеры) инверсии. При инверсиях не происходит потери генетического материала, поэтому инверсии, как правило, не влияют на фенотип носителя. Однако, если у гетерозигот по инверсиям (то есть у организма, несущего как нормальную хромосому, так и хромосому с инверсией) в процессе гаметогенеза при мейозе происходит кроссинговер в пределах инвертированного участка, то существует вероятность формирования аномальных хромосом, что в свою очередь может привести к частичной элиминации половых клеток, а также формировании гамет с несбалансированным генетическим материалом.
Более 1% человеческой популяции являются носителями перицентрической инверсии в 9 хромосоме, которую считают вариантом нормы[2]
Транслокации
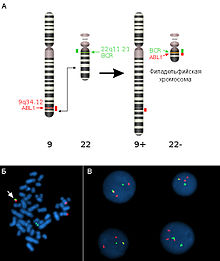
Детекция филадельфийской хромосомы при помощифлуоресцентной гибридизации in situ
Транслокации представляют собой межхромосомную перестройку, при которой происходит перенос участка одной хромосомы на другую. Отдельно выделяютреципрокные транслокации (когда две негомологичные хромосомы обмениваются участками) и Робертсоновские транслокации, или центрические слияния (при этом две негомологичные акроцентрические хромосомы объединяются в одну с утратой материала коротких плеч). Первым центрические слияния описал американец У.Робертсон (W.R.B.Robertson) в 1916 г., сравнивая кариотипы близких видов саранчовых.
Реципрокные транслокации не сопровождаются утратой генетического материала, их также называют сбалансированными транслокациями, они, как правило, не проявляются фенотипически. Однако, у носителей реципрокных транслокаций половина гамет несёт несбалансированный генетический материал, что приводит к снижению фертильности, повышенной вероятности спонтанных выкидышей и рождения детей с врождёнными аномалиями. Частота гетерозигот по реципрокным транслокациям оценивается как 1 на 600 супружеских пар. Реальный риск рождения детей с несбалансированным кариотипом определяется характером реципрокной транслокации (спецификой хромосом, вовлеченных в перестройку, размерами транслоцированных сегментов) и может достигать 40 %.
Примером реципрокной транслокации может служить транслокация типа «филадельфийская хромосома» (Ph) между хромосомами 9 и 22. В 95 % случаев именно эта мутация в гемопоэтических клетках-предшественниках является причиной хронического миелобластного лейкоза. Эту перестройку описали П.Новелл (P.Nowell) и Д.Хангерфорд (D.Hungerford) в 1960 г. и назвали в честь города в США, где оба работали. В результате этой транслокации ген ABL1 из хромосомы 9 объединяется с геном BCR хромосомы 22. Активностьнового химерного белкаприводит к нечувствительности клетки к воздействию факторов роста и вызывает её безудержное деление.
Робертсоновские транслокации являются одним из наиболее распространенных типов врождённых хромосомных аномалий у человека. По некоторым данным, их частота составляет 1:1000 новорожденных. Их носители фенотипически нормальны, однако у них существует риск самопроизвольных выкидышей и рождения детей с несбалансированным кариотипом, который существенно варьирует в зависимости от хромосом, вовлеченных в слияние, а также от пола носителя. Большинство Робертсоновских транслокаций (74 %) затрагивают хромосомы 13 и 14. В структуре обращаемости на пренатальную диагностику лидерами оказываются носители der(13;14) и der(14;21)[3]:1. Последний случай, а именно, Робертсоновская транслокация с участием хромосомы 21 приводит к так называемому «семейному» (наследуемому) синдрому Дауна.
Робертсоновские транслокации, возможно, являются причиной различий между числом хромосом у близкородственных видов. Показано, что два плеча 2-й хромосомы человека соответствуют 12 и 13 хромосомам шимпанзе. Возможно, 2-я хромосома образовалась в результате робертсоновской транслокации двух хромосом обезьяноподобного предка человека. Таким же образом объясняют тот факт, что различные виды дрозофилы имеют от 3 до 6 хромосом. Робертсоновские транслокации привели к появлению в Европе нескольких видов-двойников (хромосомные расы) у мышей группы видов Mus musculus, которые, как правило, географически изолированы друг от друга. Набор и, как правило, экспрессия генов при робертсоновских транслокациях не изменяются, поэтому виды практически неотличимы внешне. Однако они имеют разные кариотипы, а плодовитость при межвидовых скрещиваниях резко понижена.
Изохромосомы
Изохромосомы состоят из двух копий одного плеча хромосомы, соединенных центромерой таким образом, что плечи образовавшейся хромосомы представляют собой зеркальные «отражения» друг друга. В определенном смысле изохромосома представляет собой гигантскую инвертированную дупликацию размером с целое плечо и делецию другого плеча. Пациенты с 46 хромосомами, из которых одна представляет собой изохромосому, являются моносомиками по генам утраченного хромосомного плеча и трисомиками по генам, присутствующим в изохромосоме. Если изохромосома является добавочной, то данный пациент является тетрасомиком по генам, представленным в изохромосоме. В целом, чем меньше изохромосома, тем меньше генетический дисбаланс, и тем более вероятно выживание плода или ребенка с такой перестройкой. Следовательно, не удивительно, что наиболее частые из описанных случаев аутосомных изохромосом вовлекают хромосомы с маленькими плечами. Некоторые из наиболее частых участников формирования изохромосом — это короткие плечи хромосом 5, 8, 12, 18[4].
Для объяснения возникновения изохромосом можно предположить два механизма: (1) вследствие аномального поперечного разделения центромеры при делении клетки или (2) в результате неправильного слияния концов изохроматидного разрыва, образовавшегося в прицентромерной области
Дата добавления: 2016-05-30; просмотров: 3174;
Поиск по сайту
Узнать еще
- ВЛИЯНИЕ ОТКЛОНЕНИЙ В РАЗВИТИИ ПСИХИКИ РЕБЕНКА НА ВОЗНИКНОВЕНИЕ АДДИКТИВНОГО ПОВЕДЕНИЯ
- Возникновение (приобретение) и прекращение права собственности и иных вещных прав.
- Возникновение антисептики. Физико-химическая теория брожения и гниения
- Возникновение баскетбола
- Возникновение в расцвет Парфянской державы.
- ВОЗНИКНОВЕНИЕ ВИТАМИНОЛОГИИ
- Возникновение гидравлического удара
- Возникновение Голландской колониальной империи.

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
