ЖИВОТНЫХ ОРГАНИЗМОВ
МАЛОИЗУЧЕННЫЕ ТАКСОНЫ
Биологическое разнообразие живых организмов, обитающих на планете Земля, поистине огромно. Ежегодно описываются сотни и тысячи ранее неизвестных видов животных. Вполне понятно, когда по результатам экспедиций в тропические леса Амазонии или Новой Гвинеи специалисты делают первоописания ранее неизвестных видов беспозвоночных животных. Парадоксально, но в наше время возможно и первооткрытие новых типов или отделов животных и протист, – именно первооткрытие, а не описание по результатам таксономического анализа уже известных форм. Датский зоолог Р. Кристенсен даже получил в научных кругах прозвище «открывателя новых типов», причем работал он главным образом с обитателями умеренных широт.
Во второй половине прошлого века было сделано описание новых таксонов высокого ранга, информация о которых не вошла в большинство русскоязычных учебников, однако эти открытия существенно расширили представления о многообразии органического мира и его эволюции.
Если начать наш обзор с филогенетически более низкоорганизованных форм, то первым следует упомянуть трихоплакса (Trichoplax). Это единственный известный представитель типа Placozoa. Он был описан еще в 1883 г. австрийским зоологом Ф. Шульце, который обнаружил амебоидно изменяющие свою форму пластинки диаметром 1–6 мм на стенках морского аквариума в городе Грац. В 1907 г. специалист по кишечнополостным Г. Крумбах посчитал эти организмы личинками медузы Eleyteria. Лишь в 70-е годы XX века немецкий зоолог К. Грелл открыл у этих животных половое размножение.
Детальные исследования цитогистологии трихоплакса продемонстрировали уникальность организации этих животных (Рис. 1).
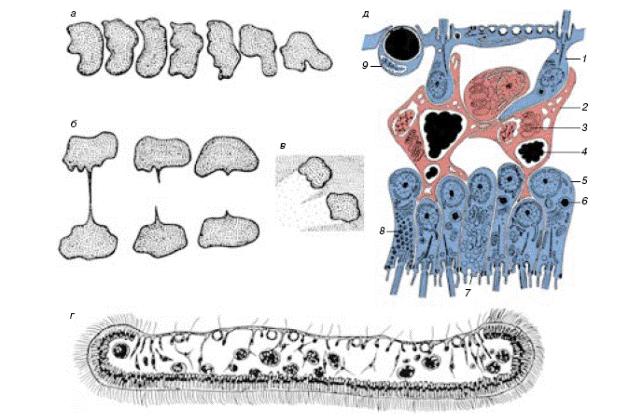
Рис. 1. Примитивный многоклеточный организм Трихоплакс: а - изменение формы тела (через промежутки 10 мин); б - деление; в - дорожки, образующиеся в культуре цианобактерий за пасущимися Трихоплаксами; г - поперечный срез пластинки Трихоплакса в световом микроскопе; д –строение Трихоплакса по данным электронной микроскопии: 1 - клетки спинного слоя, 2 - отростки волокнистых клеток, 3-митохондриальный комплекс, 4 - крупные пищеварительные вакуоли в волокнистых клетках, 5 - клетки брюшного слоя, 6 - мелкие пищеварительные вакуоли в эпителиальных клетках, 7 - слизистая клетка, 8 - железистая клетка, 9 - клетка с "блестящим шаром" (из В.В. Малахов, 2001)
Trichoplax был выделен в самостоятельный тип Placozoa. В настоящее время считается, что трихоплакс весьма близок к предкам многоклеточных. На это, помимо прочего, указывают результаты исследования рибосомальной РНК, которые подтверждают его родство с губками и примитивными кишечнополостными.
Мезозои (тип Mesozoa ) внешне схожи с Placozoa, но рассматриваются как неродственная им группа, ведущая происхождение от паразитических плоских червей. Мезозои имеют пластинчатую форму тела, его длина не превышает 1 см, чаще около 1 мм. Это полостные паразиты моллюсков и офиур, к настоящему времени описано около полусотни видов. Тело покрыто ресничным эпителием, который заключает рыхлый слой генеративных клеток. Мезозоям свойственна гетерогония – чередование партеногенетического и полового размножения. Именно исследования зародышей послужили основанием для заключения о вторично упрощенной анатомии этих животных.
Среди водных и почвенных беспозвоночных многие имеют червеобразную форму тела. Это вполне объяснимо, учитывая её универсальность и соответствие условиям обитания в плотных средах. В Линнеевской системе они были объединены в тип Vermes, затем в зависимости от основных черт анатомии их разделили на паренхиматозных (плоские черви – Plathelminthes), первичнополостных (Nemathelminthes) и вторичнополостных (кольчатые черви, эхиуриды и сипункулиды). При этом дальнейшие исследования выявили сборный характер Nemathelminthes s. l., и из состава типа были последовательно выведены сначала скребни (Acanthocephales), затем коловратки (Rotatoria), и, наконец, волосатики (Gordeimorpha, или Nematomorpha). В 1980 г. было предложено объединить волосатиков, киноринхов (Kinorhyncha) и приапулид (Priapulida) в тип Cephalorhyncha – головохоботных. Всем им присущ единый план строения: тело подразделяется (хотя бы на личиночной стадии) на 2 отдела: хоботный (интроверт) и туловищный. Интроверт уникален тем, что внутри него залегает кольцо центральной нервной системы, т.е. кольцевой мозг (таким образом, интроверт – это выворачивающаяся голова). Туловище иннервируется единичным брюшным нервным стволом.
Если киноринхи кратко описаны в классическом учебнике профессора В. А. Догеля, то приапулиды остались за его рамками. Между тем, это весьма примечательная группа морских беспозвоночных. Известно 17 рецентных, т.е. ныне существующих видов. Из них Priapulus caudatus Lamarck, 1816 имеет циркумполярное распространение, на юг проникает до Средиземного моря. Вид эврибатен, регистрируется и на мелководьях, и на глубинах до 3,5 км. Размеры тела достигают 20 см, оканчивается оно ветвистым хвостовым придатком. Животные обнаруживаются в донных илах, где они роются в поиске мягкотелых животных и съедобных органических остатков. P. caudatus отмечен в Балтийском и Белом морях, относительно обычен в Дальневосточных морях.
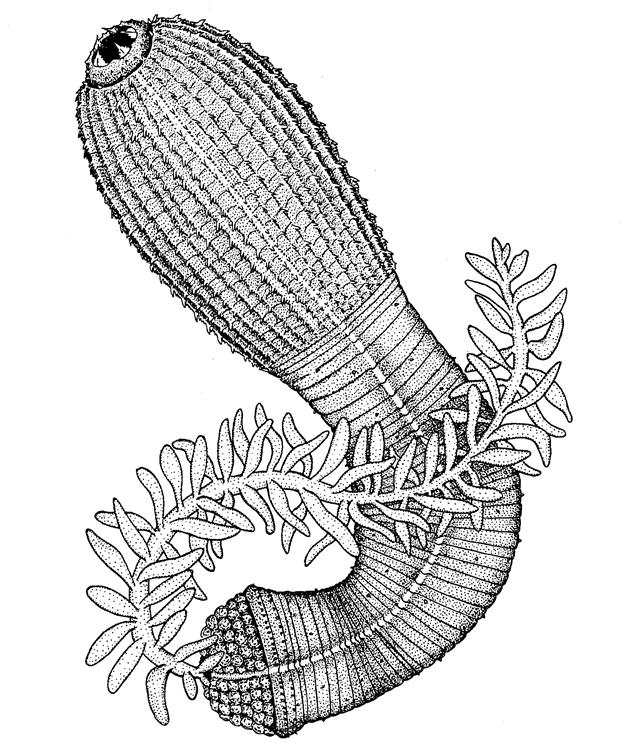
Рис. 2 Внешний облик Priapulus caudatus (из А.В. Андрианов, В.В. Малахов, 1996)
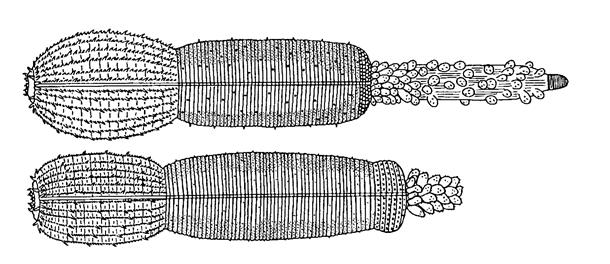 Родовое название происходит от названия греческого божества (Priap, Priapos), что связано с характерной формой тела. Внешний облик других представителей группы однообразен (Рис. 3).
Родовое название происходит от названия греческого божества (Priap, Priapos), что связано с характерной формой тела. Внешний облик других представителей группы однообразен (Рис. 3).
Рис. 3 Внешний облик приапулид Priapulus tuberculatospinosus и Priapulus abyssorum
(из А.В. Андрианов, В.В. Малахов, 1999)
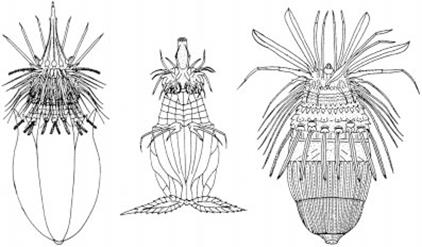 Лорициферы.В 1975 г. уже упоминавшимся датским зоологом Р. Кристенсеном был обнаружен новый вид морских червей – Nanaloricus mysticus, которых он собрал на глубине около 20 м, ныряя у стен легендарного замка Эльсинор. Собранный материал был утрачен. Повторно лорициферы были коллектированы в 1982 г. и, наконец, описаны в 1983 г. Размеры животных составляли примерно 0,5 мм, – то есть они различимы невооруженным глазом (Рис. 4).
Лорициферы.В 1975 г. уже упоминавшимся датским зоологом Р. Кристенсеном был обнаружен новый вид морских червей – Nanaloricus mysticus, которых он собрал на глубине около 20 м, ныряя у стен легендарного замка Эльсинор. Собранный материал был утрачен. Повторно лорициферы были коллектированы в 1982 г. и, наконец, описаны в 1983 г. Размеры животных составляли примерно 0,5 мм, – то есть они различимы невооруженным глазом (Рис. 4).
Рис. 3 Внешний облик лорицид (из В.В. Малахов, 2001)
Тело подразделяется на интроверт и туловище, которое заключено в кожистый панцирь – лорику. Интроверт несет набор направленных вперед стилетов, которые окружают ротовое отверстие. Затем следуют ряды чувствительных придатков – скалид, к туловищу интроверт причленяется суженным «шейным» отделом
Лорициферы обитают в промежутках между частицами морского грунта. Эти хищники, которые нападают на мелких ракообразных, прокалывают покровы тела и высасывают содержимое. Любопытно, что размеры яиц лорицифер лишь не намного меньше размеров тела взрослых червей, – разумеется, эти животные малоплодовиты. К настоящему времени известны десятки видов лорицифер, главным образом глубоководных.
Лорициферы по своей организации напоминают личинок приапулид, – предполагается, что они произошли от их неотенических личинок. Сначала лорицифер выделили в отдельный тип, сейчас преобладает мнение, что их следует объединить с приапулидами и волосатиками в тип Cephalorhyncha.
Циклиофоры, а точнее единственно известный вид Symbion pandora – также описаны Р. Кристенсеном в 1995 г. в соавторстве с П. Функом. Их обнаружили на околоротовых придатках омаров. Это сидячие организмы, питающиеся остатками пищи хозяина, – т.е. комменсалы. Максимальные размеры тела – 350 мкм, – что около порога различения человеческим глазом. На переднем конце тела расположена ротовая воронка, окруженная кольцом ресничек, на заднем – прикрепительный диск – присоска (Рис. 5). Пищеварительный тракт имеет U-образную форму как и у мшанок (Bryozoa). Размножение бесполое – внутренним почкованием. Проблему для циклиофор должны представлять регулярные линьки омаров, для преодоления затруднения циклиофоры переходят к половому размножению. Мелкие (до 80 мкм) самцы не питаются, разыскивают бесполых особей, вынашивающих самок, и прикрепляются к их покровам. Основной объем тела самцов приходится на семенные мешки. Внутри оплодотворенных самок начинается развитие личинок, которые освобождаясь, отыскивают омаров и прикрепляются к их ротовых частям, превращаясь в бесполых взрослых особей.
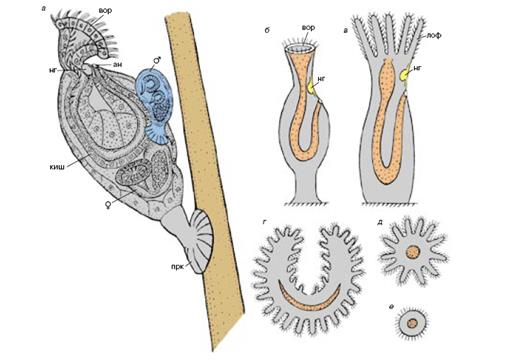
Рис. 5 Циклиофоры - миниатюрные нахлебники омаров: а - внешний вид бесполой особи
с прикрепленным к ней самцом (по: Funch, Kristensen,1995), б - план строения циклиофоры, в - план строения голоротой мшанки, г - лофофор покрыторотых мшанок, д - лофофор
голоротых мшанок, е - лофофор циклиофор, ан - анус, вор - ротовая воронка, киш - кишка, лоф - лофофор, нг - нервный ганглий, прк - прикрепительный диск, 9 - развивающаяся почка самки,♂- самец, прикрепившийся к бесполой особи в ожидании самки
(из В.В. Малахов, 2001)
В настоящее время циклиофор выделяют в самостоятельный класс в рамках типа Мшанок.
Морские маргаритки (sea daises) были впервые найдены на затонувших бревнах, поднятых с глубины несколько сот метров новозеландскими зоологами А. Бейкером, Ф. Раувом и Х. Кларком. Они отнесли их к новому классу иглокожих Concentricycloidea. Эти животные имеют уплощенное дисковидное тело диаметром до 1 см с многочисленными краевыми иглами (Рис. 6).
Радиальные каналы у эти иглокожих отсутствуют, а амбулакральные ножки располагаются по краю диска. Известно два вида морских маргариток Xyloplax medusiformis и Xyloplax turnerae. У последнего имеется рот и рудиментарный желудок. Представители первого рта и кишечника не имеют. Обращенная к субстрату поверхность тела выполнена мембранозным эндотелиальным эпителием, считается, что через него осуществляется поглощение низкомолекулярных соединений, являющихся продуктами бактериального разложения древесины.
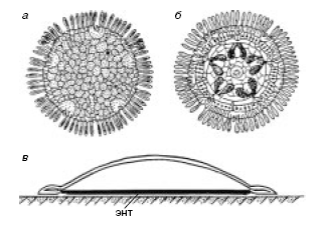
Рис. 6 Морские маргаритки: а - Xyloplax medusiformis, вид сверху, б - то же, вид снизу
(через прозрачную энтодермальную мембрану просвечивают пять пар гонад),
в - схема размещения морских маргариток на субстрате, энт - энтодермальная мембрана
(из В.В. Малахов, 2001)
Вестиментиферы представляют собой пример высокоспециализированной группы беспозвоночных, приспособившейся к обитанию в экстремальных условиях с высокими температурами и пиковыми концентрациями Н2S и сульфидов – гидротермах «черных курильщиков». Такие гидротермы обнаруживаются в рифтовых зонах. Здесь из глубин планеты по скважинам подымаются высокотемпературные газы, которые локально разогревают воду до температуры, превышающей 100º С. При повышенном давлении морских глубин закипания не происходит. Струи горячей воды бьют из трубчатых жерл, напоминающих вулканические, смешиваясь с окружающей водой, при этом горячая вода замутняется частицами образующих нерастворимых сульфидов, – это и послужило основанием для названия «черные курильщики».
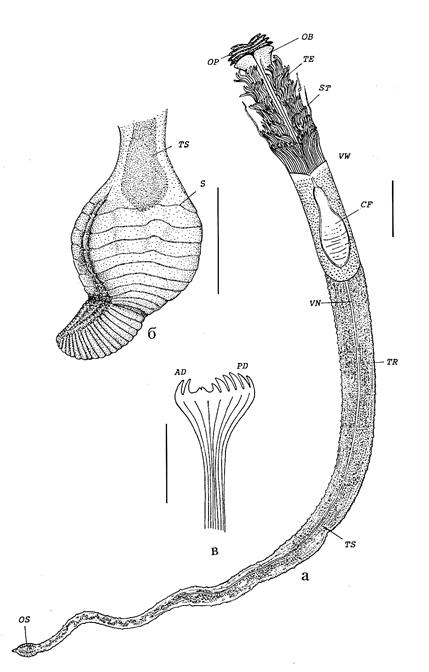
Свободная поверхность грунта непосредственно вокруг курильщика укрыта так называемыми «матами» серобактерий, выдерживающих температуры до 120º С. Также в зоне температур 30–40º С могут располагаться роговые трубки полихет – помпейских червей (Alvinella pompejana). В зоне более низких температур располагаются трубки вестиментифер. Первым описанным видом этих погонофор была Riftia pachyptila (Джонс, 1961). Размеры взрослых вестиментифер составляют от 5 см до 2,5 м. Тело заключено в белково-хитиновую трубку, обычно последние сплетаются группами десятками и сотнями особей. Из открытого конца трубки высунут султан щупалец, втягиваемый при малейшей опасности (Рис. 7).
 Характерным обитателем колоний вестиментифер является краб Bythograea, основу рациона которого составляют откушенные щупальца вестиментифер.
Характерным обитателем колоний вестиментифер является краб Bythograea, основу рациона которого составляют откушенные щупальца вестиментифер.
Вход в трубку закрывают хитиновые крышечки. Ниже зоны расположения щупалец выделяются так называемые вестиментальные крылья – боковые выросты, загибающиеся на спинную сторону тела. На брюшной стороне находится брюшное ресничное поле.
Рис. 7 Внешний облик вестиментиферы Rigdeia piscesae (из В.В. Малахов, С.В. Галкин, 1998)
В вестиментальном отделе располагаются сердце, почки и мозг. Характерно наличие гигантских аксонов, идущих от мозга к заднему концу тела, – они обеспечивают быстроту движений по втягиванию щупалец и закупорке входа в трубочку. Брюшной отдел занят трофосомой, пронизанной кровеносными сосудами. Задний отдел тела сегментирован и несет множество щетинок, заякоривающих тело вестиментиферы в трубке. Детали строения вестиментифер можно более наглядно представить из рисунка (Рис. 9).
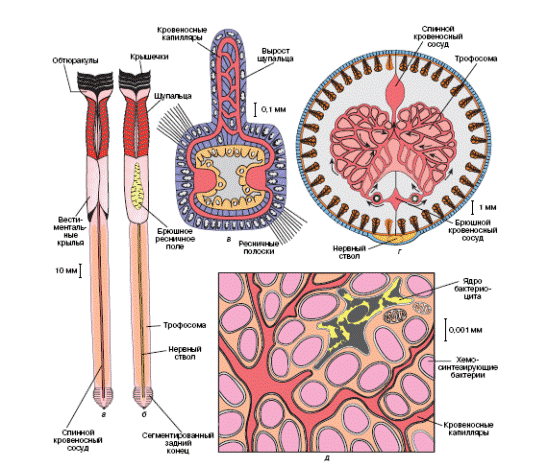
Рис. 9. Строение вестиментифер: а - внешний вид вестиментиферы Риджейа (Ridgeia) со спинной стороны; б - то же, вид с брюшной стороны; в - поперечный срез щупальца; г - поперечный срез через туловищный отдел, стрелки показывают направление движения крови; д - часть клетки-бактериоцита с пронизывающими ее кровеносными капиллярами (из В.В. Малахов, 1997)
Вакуоли клеток трофосомы содержат сероводородокисляющие бактерии Thiobacillus thiooxidans. От токсикации H2S организм защищается депонированием сульфидов в таурине. Кровеносная система вестиментифер приспособлена к транспорту как кислорода, так и сероводорода, поскольку это необходимо для обеспечения ресурсами симбионтов. Бактерии служат единственным источником органики для вестиментифер.
Впервые вестиментиферы были подняты подводным аппаратом Deepstar с глубины 1125 м континентального склона у берегов Калифорнии в 1966 г. Они были описаны М. Веббом как Lamellibranchia barkami и отнесены к новому классу погонофор.
Развитие вестиментифер осуществляется с метаморфозом: специальные исследования показали, что половые клетки и яйца свободны от бактерий – симбионтов. Личинки вестиментифер трохофорообразные, имеют хорошо развитые рот и кишечник. После короткого периода флотирования в толще воды они опускаются на дно, где заглатывают серобактерии нужного вида. Вскоре ротовое и анальное отверстия затягиваются, и кишечник претерпевает превращение в трофосому.
Исследования вестиментифер позволили пролить свет на вопрос о способе питания собственно погонофор. Погонофоры лишены рта и кишечника, но обладают специфичным органом, известным под названием «срединный канал». Оказалось, что это трофосома, населенная метанобактериями.
В противоположность болотам и грунтам пресноводных водоемов в морях метан главным образом просачивается из толщи подстилающих дно пород. Показательно, что районы обитания неглубоководных погонофор – Северное и Баренцево моря, прибрежная зона Сахалина – это места, где ведется добыча углеводородов или разведаны их запасы. Это позволяет предположить, что погонофоры могут служить биоиндикаторами месторождений газа и нефти.
Таким образом, открытие вестиментифер позволило пролить свет на невыясненные аспекты биологии погонофор в целом. Примечательно, что в гидротермальных источниках были обнаружены беспозвоночные, которых с полным основанием можно назвать «живыми ископаемыми».
В частности, это сидячие усоногие ракообразные новоописанного рода Neolepas, строение которых близко к строению ископаемых усоногих, обитавших на морских мелководьях 230–130 млн лет назад. Имеющий архаичную колпачковидную раковину брюхоногий моллюск Neomphalus представляет группу, считавшуюся вымершей около 200 млн лет назад. Карликовый морской гребешок Bathypecten имеет признаки, промежуточные между двумя ветвями линий эволюции двустворчатых моллюсков, разделившихся более 230 млн лет назад.
Описание новых для науки таксонов с одной стороны может подкрепить уже существующие гипотезы происхождения тех или иных групп организмов, с другой – разрушить ранее утвердившиеся филогенетические теории. Это справедливо как в отношении ныне существующих животных (рецентные формы), так и описания ископаемых (фоссильных) форм. Например, эпохальным событием в палеозоологии послужило нахождение и описание остатков и дериватов животных, обитавших в морях планеты в завершающий период Протерозойской эры – Венд. Благодаря этому стало возможным не только подтвердить правильность гипотезы происхождения билатерально-симметричных многоклеточных, но и прояснить филогенетические отношения между метамерными и неметамерными беспозвоночными.
Венд – заключительный период протерозойской эры (около 600 млн лет назад), когда, по всей видимости, и появились первые билатеральные животные. Он предшествовал кембрийскому периоду, когда имел место так называемый «кембрийский взрыв» – появление за эволюционно короткий промежуток практически всех мегатаксонов (типов и классов) беспозвоночных.
Фоссильные (ископаемые) остатки вендских организмов представлены отпечатками животных, не имевших твердого минерального скелета. Наиболее многочисленны формы с радиальной и лучевой симметрией, но имелись и билатерально симметричные животные.
Примечательно, что это метамерные формы, причем у некоторых сегменты располагаются со сдвигом относительно плоскости билатеральной симметрии, что характерно также для некоторых современных кораллов Ceriantharia. В отпечатках многих вендских Bilateria удается дифференцировать медиальную борозду, соответствующую центральному отделу кишечника, а также отходящие от него боковые ветви и камеры. Животные вели донный образ жизни, ротовое отверстие располагалось на брюшной стороне тела. У некоторых оно имело щелевидную форму. Предполагается, что зарастание его центральной части предопределило создание кишечной трубки. Данную гипотезу подтверждают результаты исследований экспрессии так называемых гомеобоксных генов, которые определяют развитие осевых структур, сегментацию и закладку конечностей. У кишечнополостных эти гены экспрессируются в околоротовом кольце, тогда как у позвоночных вдоль удлиненного бластопора зародышей, а затем в околоротовой и анальной зонах. Таким образом, генетические методы подтверждают предположение, что имело место удлинение первичного ротового отверстия, смыкание его посередине и превращение отверстий на его концах в дефинитивные рот и анус.
Гастральные карманы отделились от центральной части гастральной полости и превратились в целомические камеры, расположение которых определило метамерное строение тела. Сложная организация первичных Bilateria объясняет возникновение в период последующего «кембрийского взрыва» основных высокоорганизованных групп беспозвоночных, таких как олигохеты, моллюски и членистоногие. Принятие данной гипотезы меняет сложившиеся взгляды на филогению беспозвоночных животных. В частности, первичнополостные более не рассматриваются в качестве предковых для вторичнополостных (целомических) животных. В свете вышеизложенного осевая симметрия гребневиков должна считаться первичной, а сам таксон рассматриваться боковой ветвью, уклоняющейся от магистральных путей эволюции животных.
Таким образом, прогресс в исследовании фоссильных (ископаемых) и рецентных (ныне живущих) определяет существенное расширение представлений о многообразии органического мира и уточнить некоторые вопросы его эволюции.
| <== предыдущая лекция | | | следующая лекция ==> |
| Антропогенные факторы. | | | ИСТОРИЯ РАЗВИТИЯ ПСИХОЛОГИЧЕСКОЙ ПОМОЩИ ДЕТЯМ С ПРОБЛЕМАМИ В РАЗВИТИИ |
Дата добавления: 2016-07-05; просмотров: 3097;
Поиск по сайту
Узнать еще
- Адаптации организмов
- АДАПТАЦИИ ОРГАНИЗМОВ
- Адаптации организмов к условиям среды
- АДАПТАЦИЯ И ОСНОВНЫЕ СПОСОБЫ ПРИСПОСОБЛЕНИЯ ЖИВЫХ ОРГАНИЗМОВ К ЭКСТРЕМАЛЬНЫМ УСЛОВИЯМ СРЕДЫ
- Адаптация растений и животных к световому режиму.
- Активные, пассивные и избегающие адаптации организмов к неблагоприятным факторам среды
- Анаэробная энтеротоксемия животных
- АНТИГЕНЫ МИКРООРГАНИЗМОВ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
