Подкласс Бриевые (зеленые) мхи - Bryidae.
Подкласс бриевых наиболее обширный и полиморфный из всех листостебельных мхов. В нем насчитывают около 14 000 видов и более 700 родов, относящихся к 85 семействам. Это многолетние или однолетние мхи, различные по величине, от чрезвычайно мелких (высотой около 1 мм) до крупных (высотой до 50 см и более), обычно зеленые, реже красно-бурые: бурые, иногда черные.
Растут бриевые отдельными стебельками, группами, подушечками, дерновинками, ковриками или сплошными зарослями на различных субстратах — почве, коре стволов и ветвей, гнилой древесине, на содержащих известь и кислых горных породах, реже на разложившихся трупах мелких животных, на экскрементах животных, в местах различной степени увлажнения и освещения.
Распространены более широко, чем сфагновые мхи. Живут в разнообразных экологических условиях от тундры и лесотундры до степей и пустынь. Наиболее типичные местообитания бриевых мхов, где они доминируют или формируют сплошной покров, - тундра, болота и некоторые типы лесов. Каждому местообитанию свойственны свои виды. Бриевые мхи по сравнению со сфагновыми отличаются большим разнообразием строения. Органы полового размножения закладываются у одних видов на главной оси, у других - на боковых. У некоторых видов ветвление не выражено.
Политрих обыкновенный, кукушкин лен (Polytrichum commune) - это один из распространенных представителей бриевых мхов. Он растет в лесу, на полянах, окраинах болот.
Стебель гаметофита прямостоячий, неветвистый, высотой 15 см и более, густо покрыт листьями. Подземная часть его простирается в почве почти горизонтально, на ней образуются ризоиды. В центре стебля находится концентрический проводящий пучок, состоящий из вытянутых клеток, сходных с трахеидами и ситовидными трубками. Он окружен паренхимой, также выполняющей проводящую функцию. С наружной стороны паренхима граничит со склеродермой (корой). Внешний слой ее, состоящий из бесцветных клеток, называют гиалодермой.
Листья расположены по спирали. Они состоят из линейной пластинки с заостренной зубчатой верхушкой и пленчатого влагалища. На верхней стороне листа расположены ассимиляционные пластинки. Жилка с механическими и проводящими элементами расширена.
Гаметофит двудомный. Архегонии бутылковидной формы расположены на верхушке женского гаметофита, антеридии мешковидной формы - на верхушке мужского. Между архегониями и антеридиями имеются стерильные нити - парафизы. После оплодотворения из зиготы образуется спорогон, состоящий из длинной ножки и коробочки. Коробочка прямостоячая или более или менее косо расположенная, призматическая, четырех-пятиграниая, покрыта ржаво-войлочным колпачком, образующимся из стенок архегония. Коробочка состоит из урночки и крышечки. Нижняя часть урночки сужена в шейку. На границе урночки и шейки в эпидерме имеются устьица. В центре урночки расположена колонка, которая у крышечки расширяется и формирует эпифрагму - тонкостенную перегородку, закрывающую урночку. Вокруг колонки расположен спорангий в виде цилиндрического мешка, прикрепленного к стенке и колонке особыми нитевидными образованиями. Урночка имеет специальное приспособление для рассеивания спор - перистом, представляющий собой ряд зубчиков с тупыми верхушками, расположенных по краю урночки. Между зубцами, способными к гигроскопическим движениям, и эпифрагмой имеются отверстия, через которые в сухую погоду высыпаются споры. Из споры вырастает протонема в виде зеленой ветвящейся нити. На ней формируются почки, из которых со временем развиваются взрослые гаметофиты. Спорогон поднимается над перихециальными листьями или погружен в них. Коробочка обычно с перистомом, колечком и крышечкой, реже клейстокарпная. Споровый мешок пронизан колонкой (отсутствует у видов рода архидиум) и окружен, воздушной полостью. Споры большей частью округлые, гладкие или со сложной скульптурой, одноклеточные, редко многоклеточные, диаметром от 5 до 200 мкм у архидиума и некоторых родов гигаспермовых (Gigaspermaceae), чаще же 10-20 мкм.
Прорастание спор, образование протонемы и молодых растений у бриевых мхов происходит по особому типу. При прорастании спора набухает, затем эндоспорий, прорывая экзоспорий, вытягивается в нить. Эта нить ветвится, и ее ветви обладают ограниченным верхушечным ростом. Ризоиды тонкие, с бурыми стенками, косыми поперечными перегородками и с относительно немногочисленными хлоропластами или лейкопластами.
Бесполое поколение (спорофит) вырастает на гаметофите (растении полового поколения) и питается за его счет. Оно не имеет самостоятельности, развито слабо и представлено безлистным бурым стебельком, на конце которого имеется коробочка со спорами, например, как у кукушкина льна. Когда коробочка созревает, из нее высыпаются споры. Попав в благоприятные условия, из спор вырастает многоклеточная нить, из которой путем почкования развиваются несколько гаметофитов.
Когда рост гаметофита заканчивается и он оказывается подготовленным к размножению, на верхушке стебля — в середине розетки листьев — развиваются половые органы: антеридии (от греч. «антерос» — цветущий) — мужские половые органы, в которых развиваются подвижные гаметы — сперматозоиды и архегонии (от греч. «архе» — начало и «гоне» — рождение) — женские половые органы, в которых образуется неподвижная женская гамета — яйцеклетка.
В период половодья или обильных дождей мужские клетки — сперматозоиды подплывают к яйцеклетке, где и происходит их слияние. После оплодотворения образуется зигота (от греч. «зиготос» — соединенный вместе) начальная стадия развития зародыша. Из оплодотворенной зиготы на следующий год развивается коробочка на длинной безлистной ножке — спорогон. Он покрыт колпачком, в котором образуются споры.
Когда колпачок спадает, из созревшей коробочки выпадают споры и, попав в благоприятные условия, прорастают в зеленую разветвленную нить — предросток. На нем образуются почки, а из них вырастают мужские и женские экземпляры кукушкина льна. Таким образом, в жизненном цикле развития мхов происходит чередование полового и бесполого поколений.
4. Класс Печеночники. Общая характеристика. Особенности морфологии, анатомии и размножения на примере маршанции многообразной.
Общее число видов около 10 тыс. Распространелы повсеместно. Примитивность строения тела печеночников свидетельствует об их древности. Маршанция обыкновенная (Marchantia polymorpha) - типичный представитель класса. Гаметофит в виде пластинчатого таллома, длиной 10 - 12 см, ветвление верхушечное. С обеих сторон он покрыт эпидермой. Верхняя эпидерма имеет вентиляционные отверстия - устьица. Они окружены специальными клетками, расположенными в четыре ряда. Под устьицами имеются воздушные камеры. Нижняя эпидерма дает выросты - одноклеточные ризоиды и красноватые или зеленоватые чешуйки, которые иногда принимают за редуцированные листья. Под верхней эпидермой расположена ассимиляционная ткань, состоящая из вертикальных столбиков паренхимных клеток с хлоропластами. Ниже находится слой тонкостенных бесхлорофилльных паренхимных клеток. Следовательно, таллом маршанции имеет дорсивентральное строение.
На верхней стороне таллома образуются особые веточки - подставки, а на них - органы полового размножения. Маршанция - двудомное растение. На одних экземплярах подставки имеют форму сидящей на ножке девятилучевои звездочки, между лучами которой на нижней стороне расположены архегонии. На других - подставки имеют форму сидящего на ножке восьмиугольного щитка, на верхней стороне которого расположены антеридии, погруженные в антеридиальные полости. В брюшке архегония образуется яйцеклетка. После ее слияния со сперматозоидом из зиготы образуется спорогон. Он представляет собой коробочку на короткой ножке, которая прикреплена к гаметофиту гаусторией. Внутри коробочки из спорогенных клеток в результате мейоза образуются гаплоидные споры, а также элатеры - мертвые удлиненные клетки со спирально утолщенной стенкой, служащие для разрыхления массы спор, а также для выбрасывания их из коробочки. В благоприятных условиях из споры развивается предросток, или протонема. Это небольшая нить. Из ее верхушечной клетки вырастает таллом маршанции.
Вегетативное размножение осуществляется выводковыми тельцами линзовидной формы, имеющими зеленый цвет. Они образуются на верхней стороне таллома в особых корзиночках в результате деления клеток, выстилающих их дно.
.Виды маршанции имеют широкое распространение. Наиболее часто их можно встретить во влажных местах: на берегах озер и рек, по оврагам и в травянистом покрове под пологом леса.
5. Отдел Плауновидные. Общая характеристика и классификация. Время наибольшего расцвета. Происхождение листьев плауновидных (микрофиллия). Цикл воспроизведения.
Плауновидные относятся к группе наиболее древних представителей земной флоры. В конце палеозойской эры, особенно в каменноугольном периоде, плауновидные достигли своего наибольшего развития и многообразия. В то время предки современных плауновидных, называемые сигилляриями, лепидодендронами или чешуедревами, покрывали большие просторы нашей планеты. В начале мезозойской эры, в связи с изменением климата, все крупные плауновидные начинают вымирать, их вытесняют более приспособленные к новым экологическим условиям древние голосеменные.
Мелкие травянистые виды плауновидных, выживающие под пологом более конкурентоспособных голосеменных, пережили их господство и успешно приспособились к дальнейшей жизни в новых экосистемах, где преобладающими растениями стали цветковые.
Память о могучих древних плауновидных осталась в пластах каменного угля, который представляет собой окаменевшие остатки лепидодендронов-чешуедревов и других древовидных папоротников и хвощей. И, сжигая в настоящее время каменный уголь, мы в виде тепла или света электростанций получаем энергию тех солнечных лучей далеких от нас эпох, которую «законсервировали» эти растения в процессе фотосинтеза более 300 млн лет назад.
1400 видов современных плауновидных, имеющих совсем небольшое сходство со своими гигантскими предками, — это вечнозеленые многолетние травы, реже полукустарники. Очень редко они поднимают свои побеги, не имеющие камбия, выше чем на один метр. С помощью стелющихся побегов плауновидные хорошо и быстро вегетативно размножаются. При этом побеги, вырастающие на много метров в длину, создают впечатление ползучего или плывущего по почве растения. Не случайно этот большой отдел древних споровых растений получил название Плауновидные.
Побеги большинства растений этой группы покрыты спирально расположенными мелкими листьями – микрофиллами. Характерно для плауновидных также дихотомическое или вильчатое ветвление побегов. Подземные части этих растений обычно представлены корневищем с придаточными корнями и видоизмененными листьями. У некоторых видов подземные части побегов образуют своеобразный орган, несущий расположенные по спирали корни и называемый ризофором (от греч. rhiza – корень и phoros – несущий), или корненосцем. Надземные и подземные побеги плауновидных нарастают ограниченное время, поскольку со временем клетки верхушечных меристем теряют способность к делению.
Спороносные листочки (спорофиллы) плауновидных по форме, размерам и цвету могут быть похожи на вегетативные листья (трофофиллы) или в той или иной степени отличаться от них. Чередуясь со стерильными листьями, спорофиллы образуют на протяжении стебля спороносные зоны или собраны в расположенные на концах ветвей компактные образования – стробилы. У некоторых ископаемых форм плауновидных спорофиллы сидели на стебле вперемежку с вегетативными листьями, не образуя ни спороносных зон, ни стробилов.
Среди плауновидных есть равно- и разноспоровые растения, гаметофиты (заростки) которых, соответственно, либо обоеполые (т.е. на них образуются как мужские, так и женские половые клетки), либо однополые. Есть между ними и другие отличия. Заростки современных равноспоровых (представителей класса плауновых) созревают в течение 1–15 лет, достигая в длину от 2 до 20 мм. При этом они развиваются в подземных или полуподземных условиях, и питаются в значительной степени или исключительно за счет образования союза с почвенными грибами. Заростки разноспоровых плауновидных (к ним относятся все представители класса полушниковых) развиваются обычно в течение нескольких недель. Они вообще лишены хлорофилла и живут за счет питательных веществ, содержащихся в споре. Поэтому они очень мелкие и даже в зрелом виде лишь слегка выступают за пределы оболочки споры.
Половые органы, развивающиеся на заростках плауновидных, представлены антеридиями, в которых развиваются сперматозоиды с двумя или многими жгутиками, и архегониями, в которых развиваются яйцеклетки. Для того чтобы сперматозоид мог достичь яйцеклетки, необходимо наличие капельно-жидкой воды.
Надо заметить, что для того чтобы наблюдать эту фазу в цикле развития плаунов, как и других высших споровых растений, нужно проявить наблюдательность и терпение натуралиста-исследователя – разыскать крошечные заростки под травяно-кустарничковым ярусом леса очень непросто.
Оплодотворенная яйцеклетка у плауновидных сразу же, не впадая в состояние покоя, развивается в растение нового бесполого поколения – спорофит.
Все плауновидные никогда не образуют цветков — это споровые растения. Спорангии — органы спороношения — образуются в пазухах листьев или на их внутренних поверхностях. Споры могут созревать довольно долго, высыпаясь, они прорастают и дают начало гаметофитам только на влажных почвах.
Плауновидные, являясь обязательными компонентами многих экосистем, нуждаются в охране и защите. При заготовке спор, которые находили раньше широкое применение и используются сейчас в медицине и в некоторых отраслях промышленности, рекомендуется аккуратно отрезать только спороносные части растений, так называемые колоски, ни в коем случае не повреждать и не вырывать остальные части растения.
6. Начертить схему циклов воспроизведения плауна булавовидного и щитовника мужского. Укажите их сходство и различия.
У плаунов в жизненном цикле происходит чередование двух поколений, полового и бесполого.
То растение, которое мы видим в лесу– это спорофит (бесполое поколение). И размножается спорофит плауна булавовидного бесполым путем – спорами, мельчайшими пылинками без малейшего запаса питательных веществ. Размножение это очень ненадежно. Ведь из десятков тысяч спор попадет в благоприятные условия, прорастет и выживет, возможно, одна. И лишь огромное количество спор все же позволяет достичь результата.
Но попасть в благоприятные условия – это еще полдела! Некоторые споры обладают длительным периодом покоя и прорастают подземным способом. Такая спора может покрыться минеральными частичками почвы, может быть перемещена вглубь почвы водой, дождевыми червями, насекомыми. И там, в глубине, через несколько лет (от 3 до 8!) она прорастет.
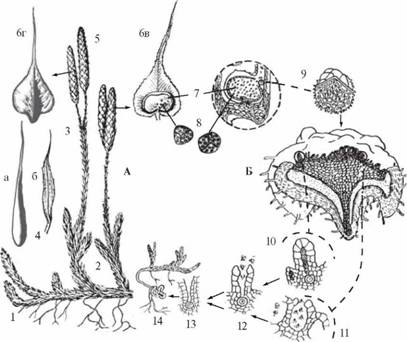 Цикл развития плауна булавовидного: А — спорофит; Б — гаметофит; 1 — ползучий побег с придаточными корнями; 2 — восходящие побеги; 3 — ножка спороносных колосков; 4 — листочки: восходящего побега (а) и ножки спороносныхколосков (б); 5 — спороносные колоски; 6 — споролистики: вид с брюшной (в) и спинной (г) сторон; 7 — спорангии; 8 — споры; 9 — прорастающая спора; 10 — архегоний; 11 — антеридий; 12 — оплодотворение; 13 — оплодотворенная яйцеклетка;
Цикл развития плауна булавовидного: А — спорофит; Б — гаметофит; 1 — ползучий побег с придаточными корнями; 2 — восходящие побеги; 3 — ножка спороносных колосков; 4 — листочки: восходящего побега (а) и ножки спороносныхколосков (б); 5 — спороносные колоски; 6 — споролистики: вид с брюшной (в) и спинной (г) сторон; 7 — спорангии; 8 — споры; 9 — прорастающая спора; 10 — архегоний; 11 — антеридий; 12 — оплодотворение; 13 — оплодотворенная яйцеклетка;
Дата добавления: 2016-05-30; просмотров: 7486;
Поиск по сайту
Узнать еще
- ВЗАИМООТНОШЕНИЯ МЕЖДУ УПРАВЛЕНИЕМ «СУРГУТНЕФТЕПРОМХИМ» И СТРУКТУРНЫМИ ПОДРАЗДЕЛЕНИЯМИ ОАО «СУРГУТНЕФТЕГАЗ» ПРИ ПРОВЕДЕНИИ РАБОТ ПО ПОВЫШЕНИЮ НЕФТЕОТДАЧИ ПЛАСТОВ
- Класс Печеночные мхи, особенности строения вегетативного тела, размножения, цикла развития на примере маршанции.
- Классы имуноглобулинов. Известно пять классов иммуноглобулинов, обозначаемых как IgG, IgM, IGA, IGD, IgE. У отдельных классов описаны подклассы.
- Подкласс AMMONOIDEA АММОНОИДЕИ
- Подкласс CLADISTIA - Кладистрии
- Подкласс Pulmonata. Легочные C-Q
- Подкласс кистеперые

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
