В интеграции метаболизма
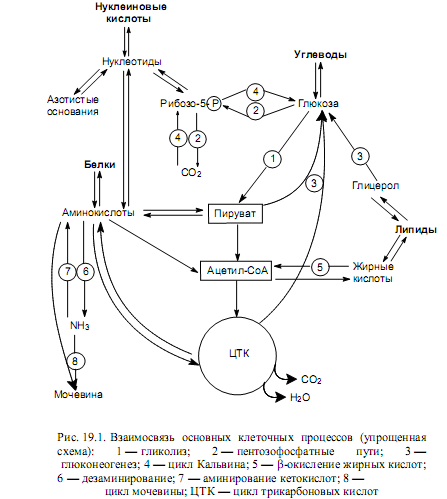
Пируват. Эта трехуглеродная a-кетокислота связывает между собой гликолиз и глюконеогенез, а также обмен углеводов с обменом липидов, белков, изопреноидов, кетоновых тел (рис. 19.1).
Пируват образуется в клетках в основном при катаболизме гексоз, при окислении лактата, который накапливается в мышцах при молочном брожении, а также при дезаминировании аланина. Процессы катаболизма гексоз охарактеризованы в главе 9. Молочнокислое брожение —основной метаболический процесс (глава 10), протекающий в клетках активно работающих мышц человека и животных при недостаточном снабжении их кислородом. При этом образование лактата позволяет организму быстро регенерировать NAD+, необходимый для запасания АТР в гликолизе (процесс сокращения и расслабления мышечных волокон требует гидролиза АТР).
Накапливающийся в мышцах лактат поступает в кровь и переносится в печень, где окисляется снова в пируват. В ходе глюконеогенеза пируват может превращаться в глюкозу. Эта последовательность реакций известна как цикл Кори.
Аланин образуется из пирувата в реакциях трансаминирования (рис. 16.8) с участием аминокислот, образованных в ходе протеолиза белков. Аланин с током крови поступает в печень, где снова превращается в пируват, выполняя роль переносчика аммонийного азота, который в печени включается в цикл мочевины. Кроме этого, образующийся из пирувата аланин может включаться в состав пептидов.
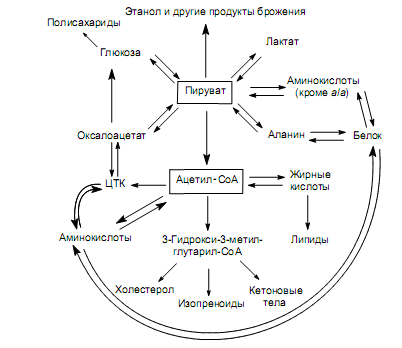
Превращения пировиноградной кислоты (рис. 19.2) связаны также с реакциями карбоксилирования и декарбоксилирования. В первом процессе, идущем в митохондриях, образуется оксалоацетат, который может включаться в ЦТК (одна из важнейших анаплеротических реакций) либо преобразовываться в фосфоенолпируват, а затем — в глюкозу (глюконеогенез).
Во втором случае пируват подвергается окислительному декарбоксилированию с образованием ацетил-СоА, который также является ключевым метаболитом.
Еще одной важной частью превращений пирувата являются различного рода брожения, в которых пировиноградная кислота служит субстратом.
Ацетил-СоА. Основными источниками этого ключевого промежуточного соединения служат пируват и жирные кислоты (процессы окислительного декарбоксилирования и b-окисления соответственно). Кроме этого, ацетил-СоА образуется из кетогенных аминокислот при расщеплении их углеродных скелетов.
Судьба ацетил-СоА, как и других ключевых метаболитов, зависит от потребностей клетки (организма): он может быть полностью окислен в цикле трикарбоновых кислот, являясь его субстратом; может подвергаться последовательному конденсированию с образованием 3-гидрокси-3-метил-глутарил-СоА — предшественника холестерола, терпеновых соединений (изопреноидов), кетоновых тел (ацетоуксусная, 3-гидроксимасляная кислоты и ацетон); а также может превращаться в жирные кислоты (рис. 19.2). Уже отмечалось, что в организме млекопитающих ацетил-СоА не способен превращаться в углеводы. В то же время в клетках растений и микроорганизмов возможен синтез предшественников глюконеогенеза, а значит, и углеводов из ацетил-СоА (рис. 11.7).
Кроме перечисленных реакций, ацетил-СоА принимает участие в процессах синтеза аминокислот, в частности аргинина, лейцина, лизина (в клетках грибов), цистеина (у некоторых микроорганизмов), а также является субстратом для маслянокислого и ацетонобутилового брожения.
Участие основных промежуточных соединений ЦТК (оксалоацетата, a-кетоглутарата) в интеграции метаболизма нетрудно проследить на схемах, представленных на рис. 19.1 и 19.2.
Подводя итог сказанному, следует еще раз подчеркнуть, что все метаболические процессы, протекающие в клетке (организме), связаны между собой с помощью промежуточных метаболитов, которые являются продуктами одних метаболических путей и субстратами других. Такая взаимосвязь и взаимозависимость метаболических процессов позволяет клетке (организму) координировать свои возможности с потребностями и быстро «настраивать» уровень обмена веществ в соответствии с меняющимися условиями существования. Конечно, этот тонкий механизм был бы невозможен без регуляции метаболических путей.
Дата добавления: 2016-05-30; просмотров: 2458;
Поиск по сайту
Узнать еще
- В. ИЗМЕНЕНИЕ ГОРМОНАЛЬНОГО СТАТУСА И МЕТАБОЛИЗМА ПРИ ГОЛОДАНИИ
- Вопрос 4. Питание и особенности метаболизма бактерий
- Два подхода к интеграции CAD- и САМ-систем
- ЗАКОНОМЕРНОСТИ МЕТАБОЛИЗМА
- ИЗМЕНЕНИЯ ВНУТРИКЛЕТОЧНОГО МЕТАБОЛИЗМА ПРИ ПОВРЕЖДЕНИИ
- Изменения метаболизма в печени в абсорбтивном периоде
- Изменения метаболизма в послеоперационном периоде

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
