Применяемые в селекции и пищевой промышленности
Микроорганизмы существенно отличаются друг от друга по морфологии, размерам клеток, отношению к кислороду, по потребностям к ростовым факторам, способности ассимилировать разные компоненты субстрата и т. д.
Из более 100000 известных видов микроорганизмов в промышленности используют относительно мало – около 100 видов. Они должны соответствовать следующим требованиям:
1. Расти на дешёвых и доступных субстратах;
2. Обладать высокой скоростью роста биомассы и давать высокую продуктивность целевого продукта при экономичном потреблении питательного субстрата;
3. Проявлять направленную биосинтетическую активность при минимальном образовании побочных продуктов (рис. 2.1);
|
|
| |||||||
| |||||||||
| |||||||||
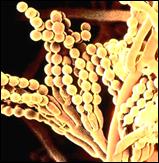
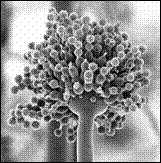


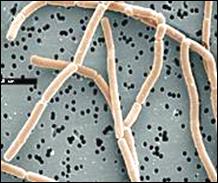
Рис. 2.1. Микроорганизмы, используемые в промышленном синтезе
различных соединений:
А – Аcetobacter aceti; Б – Aspergillus niger; В – Penicillium chrysogenum; Г – Lactobacillus delbruecki; Д – Производство левана при ферментации среды, содержащей Zymomonas mobilis
4. Быть генетически однородными, стабильными в отношении продуктивности и требований к питательному субстрату, а также условиям культивирования;
5. Быть устойчивыми к фагам и другой посторонней микрофлоре;
6. Быть безвредными (не обладать патогенными свойствами) для людей и окружающей среды;
7. Желательно, чтобы продуценты были термофильными и ацидофильными, так как в этом случае легче предохранить ферментируемый субстрат от инвазии посторонней микрофлоры;
8. Целевой продукт биосинтеза должен иметь экономическую и народнохозяйственную ценность и легко выделяться из сброженного субстрата.
Возрастающий интерес представляют анаэробные микроорганизмы, поскольку при культивировании не требуют энергоёмких аэрирующих устройств.
Сверхсинтез, т. е. способность микроорганизма синтезировать определённый продукт в количествах, превосходящих его физиологические потребности, довольно часто встречается в природе. Нередко тот или иной продукт обмена веществ (органические кислоты, спирты, антибактериальные вещества), выделяемый микроорганизмом в окружающую среду, является токсичным для других видов и служит продуценту как средство защиты обитаемого пространства или как резерв питательного вещества. Микроорганизмы с такими свойствами первыми были привлечены к хозяйственной деятельности человека в тысячелетней давности и был проведён стихийный отбор наиболее продуктивных форм. Сейчас такие природные штаммы микроорганизмов, иногда после сознательного отбора, применяют для производства микробной биомассы (микробного белка) в качестве бактериальных азотных удобрений, биопестицидов, в производстве пищевых продуктов и в других отраслях народного хозяйства. Однако основной контингент промышленных микроорганизмов представлен искусственно селекционными штаммами.
В настоящее время в промышленности применяют три вида штаммов:
1. Природные штаммы, нередко улучшенные естественным или искусственным отбором;
2. Штаммы, изменённые в результате индуцированных мутаций;
3. Штаммы культуры, полученные методами генной или клеточной инженерии.
Принципы селекции организмов. Начиная сознательную селекцию микроорганизмов, человек ставил целью создать промышленные организмы с необычными для диких микробов свойствами. Методологически эту цель человек решает двумя путями:
1. Коррекцией генетической информации микробной клетки, исключая не желаемые для промышленного синтеза свойства и усиливая нужные признаки.
2. Индукцией совершенно новой информации в генетической программе клетки.
Для этого необходимо также решить следующие задачи:
1. Существенно увеличить продуктивность, свойственную данному виду микробов и его продукту обмена веществ;
2. Генетически запрограммировать биосинтез таких веществ, которые несвойственны данному виду или даже несвойственны микробному миру.
В отличие от селекции культурных растений и домашних животных, имеющей тысячелетнюю историю и богатый опыт, целенаправленный отбор и селекция микроорганизмов начались только после узнавания микромира и развивались параллельно с достижениями генетики как научной дисциплины.
Селекционируя любой живой организм, человек опирается на естественные движущие силы эволюции – наследственные изменения и отбор положительных экземпляров. Однако микроорганизмы, как объект селекции, имеют ряд особенностей:
1. Выращивание микробной культуры из одной клетки – обычный для микробной селекции приём – приводит к тому, что в руках селекционера в качестве исходного материала селекции всегда оказывается особь клона (имеющего генетическую однообразность). С другой стороны, клон микроорганизмов быстро достигает такой численности, что за счёт естественных мутаций превращается в популяцию – совокупность клеток с разными генотипами;
2. Большинство микроорганизмов гаплоидные, с одним экземпляром хромосом, поэтому у них нет скрытой изменчивости, являющейся основой селекции высших организмов;
3. У большинства микроорганизмов, имеющих промышленное значение, до сих пор не известна способность к гибридизации (половому размножению). Это означает, что селекцию клеток можно вести только вегетативным путём;
4. Микроорганизмы характеризуются исключительно быстрой сменой поколений, поэтому возможностей для отбора положительных экземпляров у селекционера микробиолога значительно больше. Оценку выбранного микроорганизма можно провести за считанные дни выращивания в отличие от макроорганизмов, где результаты работы видны через несколько лет;
5. Селекционер микроорганизмов имеет огромное число индивидуумов для отбора, что принципиально расширяет его возможности, но для оценки продуктивности каждого клона требуется трудоёмкая работа.
Культуру, подлежащую селекции, можно выбрать из собранных в коллекции культур (музейные культуры); можно использовать известные промышленные продуценты; можно изолировать микробы из природных субстратов.
Представления о биохимии и физиологии микроорганизмов ориентируют селекционера на определённые группы микроорганизмов с наиболее вероятным потенциалом сверхсинтеза интересующего вещества. Продуценты антибиотиков, главным образом, встречаются среди грибов аскомицетов и актиномицетов, сверхсинтез аминокислот легче получить у коринебактерий, внеклеточные ферменты часто синтезируют дрожжи. Главные этапы селекции микроорганизмов отражены на рисунке 2.2.
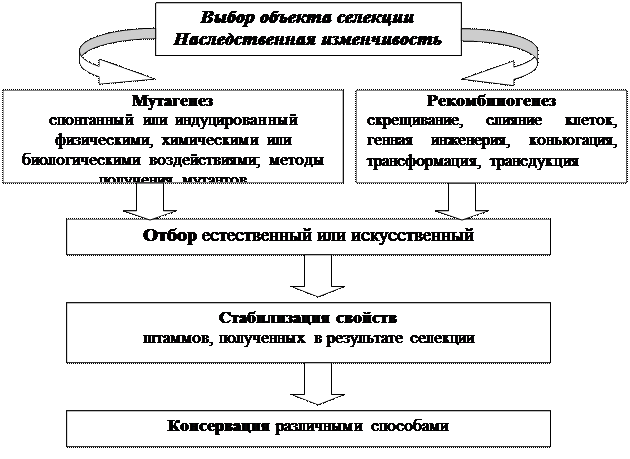
Рис. 2.2. Схема селекции микроорганизмов
Таблица 2.1. Группы методов клеточной технологии
| 1 группа: облегчение и ускорение селекционного процесса | 2 группа: создание генетического разнообразия и скрининга генотипов с важными признаками |
| Оплодотворение in vitro | Использование сомаклональных вариаций и получение индуцированных мутантов на клеточном уровне |
| Культура незрелых гибридных семяпочек и зародышей (эмбриокультура) | Клеточная селекция |
| Регенерация растений из тканей летальных гибридов | Гибридизация соматических клеток |
| Экспериментальная гаплоидия | Перенос чужеродных цитоплазматических генов |
| Клональное микроразмножение новых сортов, гибридов, линий (включая создание искусственных семян) | Перенос чужеродной генетической информации различного происхождения |
| Криосохранение генофонда | Адресный перенос ядерных генов |
Клеточная технология в селекции растений.Ускорение и облегчение селекционного процесса, а также создание растений с новыми качествами – это направления, которые достаточно успешно развиваются с помощью биотехнологических методов клеточной инженерии, культуры клеток и тканей.
Две группы методов, данной технологии, представлены в табл. 2.1.
Первые три, из указанных методов, стали традиционными, другие находятся на начальных этапах разработки. Наконец, есть такие методы, которые явно вышли из ранга вспомогательных, ускоряющих селекцию технологий. К ним можно отнести криосохранение генофонда – технологию, в настоящий момент приобретшую экологическую направленность; или клональное микроразмножение растений, тесно связанное с проблемой их оздоровления от вирусных и других инфекций.
Методы клеточной инженерии позволяют значительно ускорить традиционный процесс селекции. Биотехнология позволяет также скрещивать растения, которые в обычных условиях не скрещиваются рис. 2.2. – 2.6.
Одна из наиболее важных технологий, из перечисленных выше – оплодотворение in vitro, помогает предотвратить прогамную несовместимость, которая может быть вызвана рядом следующих причин:
1. Генетически детерминированное (определённое) несоответствие секрета рыльца материнского растения и пыльцы отцовского, которое тормозит рост пыльцевых трубок на рыльце пестика;
2. Несоответствие длины столбика пестика и пыльцевой трубки, в результате чего пыльцевая трубка не достигает семяпочки (гетеростилия);
3. Тканевая несовместимость партнёров, приводящая к остановке роста пыльцевой трубки в любой момент её прорастания, от рыльца пестика до микропиле семяпочки (гаметофитный тип несовместимости).
Преодоление прогамной несовместимости возможно благодаря выращиванию в стерильных условиях изолированной завязи с нанесённой на неё пыльцой или изолированных кусочков плаценты с семяпочками, рядом с которыми, или непосредственно на ткани которых, культивируется пыльца.
Значительным препятствием в селекции служит постгамная несовместимость, вызванная разновременным развитием зародыша и эндосперма при отдалённой гибридизации. В результате образуются невсхожие щуплые семена. Получить растение из таких семян можно только при использовании метода эмбриокультуры, т.е. выращивания изолированного зародыша на искусственной питательной среде in vitro. Метод эмбриокультуры широко применяют при межвидовой гибридизации овощных растений, для микроразмножения ценных гибридов, для клеточной селекции.
Большое значение имеет создание гаплоидов, позволяющее ускорить процесс селекции в 2-3 раза. Использование гаплоидных клеток и гаплоидных растений способствует обнаружению экспрессии введённого в клетку генома, редких рекомбинаций, рецессивных мутаций, которые в диплоидных растениях, как правило, маскируются доминантными генами. Из гаплоидных клеток можно выделить протопласты; сливаясь, они образуют гибридные клетки и растения с диплоидным числом хромосом. Обрабатывая гаплоидные клетки колхицином, можно добиться удвоения числа хромосом и получить диплоидные гомозиготные растения.
Методы селекции растений.
Методы создания дальнородственных гибридов.
| 1. Изолированные завязи | 2. Слияние протопластов |
Выращивание в стерильных условиях семяпочек одних видов растений рядом с пыльцой растений других видов (рис. 2.3; 2.4).
 Рис. 2.3. Отбор
пыльцы
Рис. 2.3. Отбор
пыльцы
 Рис. 2.4. Изолированная завязь
Рис. 2.4. Изолированная завязь
| Соматическая гибридизация. Благодаря этому методу можно осуществить любые скрещивания. Действием препаратов, разрушающих клеточную стенку, получают протопласты (рис. 2.5).
 Рис. 2.5. Протопласты
Слияние протопластов с образованием гибридных клеток происходит в присутствии полиэтиленгликоля. Продукты слияния протопластов культивируют на питательных средах с осмотическими стабилизаторами, при этом они образуют новую клеточную стенку, осуществляют ряд последовательных делений и превращаются в колонии каллусных клеток (рис. 2.6.).
Рис. 2.5. Протопласты
Слияние протопластов с образованием гибридных клеток происходит в присутствии полиэтиленгликоля. Продукты слияния протопластов культивируют на питательных средах с осмотическими стабилизаторами, при этом они образуют новую клеточную стенку, осуществляют ряд последовательных делений и превращаются в колонии каллусных клеток (рис. 2.6.).
 Рис. 2.6. Колонии каллусных клеток
После этого их переносят на среду для регенерации, где происходит образование зачатков стеблей, корней, а затем и регенерация химерного растения.
Рис. 2.6. Колонии каллусных клеток
После этого их переносят на среду для регенерации, где происходит образование зачатков стеблей, корней, а затем и регенерация химерного растения.
|
Всё это значительно облегчает выявление и стабилизацию необходимых признаков. Кроме селекции гаплоиды применяются также в генно-инженерных исследованиях. Впервые возможность получения спонтанных гаплоидов при аномальном развитии пыльников и пыльцы была показана в 1984 году С. Гуха и С. Магешвари. В настоящее время в культуре гаплоидные растения получают из изолированных пыльников (андрогенез), изолированных семяпочек (гиногенез); из гибридного зародыша, у которого в результате несовместимости потеряны отцовские хромосомы (партеногенез). Новые сорта ячменя – Исток и Одесский 15 – были выведены благодаря комбинации партеногенетического метода с культурой изолированных зародышей за 4 года вместо 10-12 лет, необходимых при обычной селекции.
Дата добавления: 2016-05-28; просмотров: 3452;
Поиск по сайту
Узнать еще
- I. Крахмал и гликоген. Роль крахмала в пищевой промышленности.
- VI. Пектиновые вещества и их роль в пищевой промышленности.
- VII. Сигналы, применяемые при маневровой работе
- VIII. Сигналы, применяемые для обозначения поездов, локомотивов и другого железнодорожного подвижного состава
- Авторы работ по лесной генетике и селекции, осуществленных в 50-х годах
- Адсорбция на твердых поверхностях и ее применение в пищевой промышленности
- АМ, применяемые в нефтяной и газовой промышленности
- Бескерновое бурение и применяемые долота

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
