Врожденный пусковой механизм
Ранние этологи (Uexkull, 1934; Lorenz, 1935) полагали, что животные нередко реагируют инстинктивно на специфические раздражители, порой достаточно сложные. Такие раздражители стали называть сигнальными раздражителями (знаковыми стимулами). Пример реакции на такой стимул изображен на рис. 20.1. Сигнальный раздражитель представляет собой какую-то часть общей структуры стимула, и эта часть может быть относительно простой. Например, у самца трехиглой колюшки в брачный период брюшко становится ярко-красным. Это и есть сигнальный раздражитель, который вызывает агрессию со стороны другого самца, охраняющего свою территорию. Как можно видеть на рис. 20.2, для про-
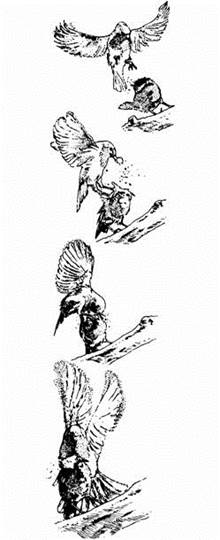
| Рис. 20.1. Малиновка атакует модель птицы. Красная грудка модели - это сигнальный раздражитель. (Рисунок по фотографии из работы Sparks, 1982.) |
воцирования агрессии достаточно и грубой модели, лишь бы нижняя ее часть была окрашена в красный цвет. Напротив, только что убитый самец колюшки, у которого брюшко не красное, уже не может вызвать атаки со стороны других самцов. Таким образом, многие другие признаки самца колюшки, по-видимому, игнорируются другими самцами. Красный цвет становится гораздо более эффективным, если им окрашена нижняя часть модели.
Именно такие сигнально-структурные взаимоотношения составляют общую черту сигнальных раздражителей, однако в случае с колюшкой это, видимо, не столь существенно. Тинберген (Tinbergen, 1953) описывает свои опыты по изучению территориального поведения, которые он проводил на самцах колюшки в аквариуме, помещенном на окне. Всякий раз, когда мимо окна проезжал красный почтовый фургон, рыбы тут же пытались «атаковать» его, как-будто это был самец-соперник. Не так давно сотрудники телевизионной компании Би-би-си имели возможность повторить эти эксперименты, когда снимали для телевидения исторический документальный фильм (Sparks, 1982).
Основываясь на том, что такого рода стимулы вызывают совершенно определенные реакции, ранние этологи предположили наличие в организме животного какого-то внутреннего механизма узнавания сигнальных раздражителей. Этот предполагаемый механизм стали называть врожденным пусковым механизмом (ВПМ) (Lorenz, 1950; Tinbergen, 1950). Сформулированная концепция имеет три важных аспекта. Во-первых, подразумевается, что этот механизм врожденный, а это означает, что и распознавание сигнального раздражителя, и реализация ответной реакции на него тоже врожденны и составляют видовую особенность животного. Однако, как мы уже отмечали в гл. 3, эти ранние представления этологов о врожденности несколько отличаются от тех, что приняты в настоящее время. Во-вторых, имеется в виду, что ВПМ играет роль высвободителя (спускателя) ответа на сигнальный раздражитель. При этом полагают, что ВПМ сдерживает заключенную в организме специфическую (для данного действия) энергию, или побуждение, до тех пор, пока животное не опознает соответствующий сигнальный
раздражитель, в ответ на который и высвободится энергия в виде соответствующего поведения. Этот аспект ВПМ был настолько существенным, что сигнальные раздражители часто называли «релизерами» (от англ. release - высвобождать, спускать). И наконец, в-третьих, считают, что этот поведенческий ответ, высвобожденный с помощью ВПМ, стереотипен и является частью врожденного репертуара, представленного комплексами фиксированных действий. Последние, как первоначально предполагал Лоренц (Lorenz, 1932), - это определенные действия, характеризующиеся достаточно жесткой координацией, что-то вроде рефлексов. Считалось, что они врожденны и видоспецифичны.
Лоренц проводит различие между комплексами фиксированных действий и рефлексами, указывая при этом на несколько пунктов. Прежде всего комплексы фиксированных действий могут быть высвобождены в ответ на различные раздражители, тогда как рефлексы вызываются лишь специфическими стимулами. Кроме того, для выполнения комплексов фиксированных действий животные должны испытывать мотивацию, тогда как для рефлекторных действий этого не нужно. И наконец, комплексы фиксированных действий могут проявиться и в отсутствие внешних раздражителей - тогда их называют холостыми действиями. Некоторые из этих пунктов сегодня представляются спорными. Например, рефлекс испуга можно вызвать различными стимулами. Помимо этого, сейчас известно, что многие комплексы фиксированных действий генерируются как вполне предсказуемые, без участия обратной связи, чем и определяется стереотипность их проявления.
Типичный пример анализа наблюдений с точки зрения концепции ВПМ мы находим у Берендса (Baerends, 1950).
«Мы располагаем важными данными в пользу представлений о том, что всякому пусковому механизму соответствует свой собственный сигнальный раздражитель. Например, роющая оса Ammophyla adriaansei, которая ловит гусениц и притаскивает их в гнездо в качестве пищи для своих личинок, может реагировать на гусеницу различным образом в зависимости от того, какой из ее инстинктов в этот момент будет активирован. Во время охоты она ловит гусеницу и парализует ее. Когда же гусеница оказывается около гнездового отверстия сразу после того, как оса открыла его, оса затаскивает ее внутрь. Но когда гусеница лежит близко к гнезду и оса заделывает вход в него, гусеница может быть использована как строительный материал. И, наконец, если мы положим ее в «гнездовую шахту» в то время, когда оса строит гнездо, она выбросит гусеницу точно так же, как она поступает с другим мешающим ей предметом, например с кусочком корешка растения. Таким образом, один и тот же объект в зависимости от состояния животного вызывает у него различные ответы. Однако во всех этих ситуациях от гусеницы постоянно идут зрительные и химические сигналы к чувствительным органам осы, где они всегда трансформируются в импульсы. Отсюда следует, что именно активированный в данный момент инстинкт осы определяет, какие из этих импульсов будут на каком-то этапе

|
| Рис. 20.2. Модели самца трехиглой колюшки, которые вызывают нападения со стороны самца, охраняющего свою территорию. Вверху - мертвый самец, утративший брачный наряд. Изображенные под ним четыре модели очень примитивны, но их красная нижняя часть оказывается достаточно эффективным сигнальным раздражителем, чтобы вызвать нападение. (По Tinbergen, 1951.) |

Рис. 20.3. Серый гусь возвращает в гнездо выкатившееся яйцо. (Из The Oxford Companion to Animal Behaviour. 1981.)
их пути блокированы, а какие смогут пройти по еще неизвестному нам пути в нервной системе, чтобы в конце концов стимулировать главный моторный центр данной реакции. Имеются данные о том, что в каждом случае эффективными оказываются различные стимулы. Весьма вероятно, что во время охоты оса воспринимает присутствие гусеницы по запаху, но когда она оставляет гусеницу при перетаскивании ее к гнезду, то для ее отыскания она использует в первую очередь зрительные сигналы».
Здесь мы познакомились с некоторыми элементами общего феномена фильтрации стимулов, о котором говорилось в гл. 12.5. Показав, что животные избирательно реагируют на сложные стимулы, этологи внесли большой вклад в понимание того, как осуществляется перцепция у животных. Вместе с тем их концепция ВПМне лишена целого ряда уязвимых мест. Так, например, Хайнд (Hinde, 1966) отмечает, что при использовании термина ВПМчасто подразумевается, что этот механизм специфичен для какой-то определенной реакции, но доказательства его существования основаны на изучении только этой реакции. Однако вполне вероятно, что особый сигнальный раздражитель может быть связан не только с одним аспектом поведения. Кроме того, избирательную реактивность животного нельзя связывать исключительно с теми стимулами, которые «запускают» реакцию. Она может проявляться и в ответ на аспекты стимула, которые важны для ориентации поведения. И наконец, механизм избирательности не обязательно должен быть исключительно врожденным; на него может влиять и научение. Как было показано в гл. 12.5, представление о сигнальном раздражителе не очень далеко отстоит от представлений об избирательном внимании - концепции, которая зародилась при исследовании процесса научения у животных.
Среди первых этологов были и такие, которые хорошо знали о существовании этих проблем. Так, Тинберген (1951) особо подчеркивал, что имеются существенные различия между пусковым (высвобождающим) и направляющим аспектами стимула. Он демонстрировал это различие на примере поведения самки серого гуся, которая возвращает в гнездо выкатившееся яйцо (см. рис. 20.3); это поведение в тридцатые годы они изучали вместе с Лоренцом.
Дата добавления: 2016-06-18; просмотров: 3401;
Поиск по сайту
Узнать еще
- CIH — газораспределительный механизм с распределительным валом в головке блока цилиндров
- III. Механизмы регуляции количества ферментов
- TActionList - механизм действий
- Аграрная реформа 1861г. и механизм ее реализации на Беларуси
- Антихолинэстеразная теория механизма действия ФОС
- Аппаратный механизм преобразования виртуального адреса в физический
- Афферентный механизм произвольных движений afferentis — «приносящий»
- Аэробный механизм ресинтеза АТФ

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
