Постав и размер ушей
На основании исследований Н. Ильина (1932) показано, что полустоячие коллиподобные (НаНа) уши имеют полное доминирование над висячими (НН) и стоячими (hh). Висячее ухо имеет неполное
Существует крайне мало данных о наследовании формы лапы и прибылых (первых) пальцев. Э. Хамфри и Л. Уорнер (1934) полагают, что "кошачья" форма лапы у собак имеет полное или частичное доминирование над "заячьей". Что касается наследования прибылых пальцев, то мнения специалистов по этому вопросу расходятся. Л. Уитни (1971) считает, что прибылые пальцы на задних конечностях доминируют над их отсутствием, что подтверждено для немецких овчарок и бракков. Однако более ранние работы К. Килера и X. Тримбла (1938), проводивших скрещивания далматиков с колли, не дают оснований для такого вывода.
М. Уиллис (2000) приводит следующие данные о наследовании формы и длины хвоста у собак:
• Полное или неполное доминирование короткого хвоста (Т) над хвостом нормальной длины (t) с летальным действием в доминантном гомозиготном состоянии (М. Берне и М. Фрэзер, 1966).
• Короткохвостость американских кокер-спаниелей наследуется как простой рецессивный признак и не имеет ничего общего с куцехвостостью (Т. Паллиг, 1953, 1957). К. Стокард и другие (1936) приводят сходные данные о доминировании штопорообразных длинных хвостов у английских и французских бульдогов над короткими.
• Существует аутосомно-доминантный ген с неполной пенетрант-ностью, обусловливающий короткий и толстый хвост (И. Залер, 1954; Р. Кертис и другие, 1964). Отбор по этому признаку может привести к сокращению хвостовых позвонков, а при инбридинге — к появлению особей с расщелиной позвоночника.
Собрано большое число данных о наследовании количества зубов, но окончательного вывода по этому поводу до сих пор не сделано. При вязках немецких овчарок с полным набором зубов Э. Хамфри и Л. Уорнер (1934) получили 202 щенка, из которых у 88 отсутствовал хотя бы один зуб. При спаривании неполнозубых собак в потомстве из 38 щенков 21 имели полный набор зубов. Наши исследования (М. Гладких, Е. Федорина, 2000) показали, что в потомстве кобеля немецкой овчарки, у которого отсутствовали премоляры (Р2), у 50% его сыновей обнаружен этот же недостаток.
Обоняние
В серии работ (Н. Чекунчикова, В. Лавровский, 1999; М. Степура, Е. Горелова, М. Гладких, 2001) показано, что порог обонятельной чувствительности собак разных пород достоверно не различается и лежит в пределах от 1 • 10~4 мл/см2 до 4 • 10~4 мл/см2. Выявлен значительный уровень внутрипородного разнообразия по пороговой чувствительности, коэффициенты вариации колеблются от 34,1% у группы бойцовых собак до 56,7% у специально выведенной для поисковой работы группы шакало-собачьих гибридов.
Существует мнение, что на обоняние оказывает влияние половая принадлежность собак. Так в работах А. Мызниковой (по Крушине-кому, 1993) отмечается, что суки в тестах по обнаружению запахового следа органических кислот оказались более чувствительными, чем кобели. Однако проводимые Н. Чекунчиковой (1999) исследования не подтвердили этого, как и достоверных различий между животными, подготовленными для разных видов служб, например для патрульно-постовой службы или же специально обученных поиску людей.
Выявлено, что повторяемость данного признака, как индивидуальная, так и групповая, практически равна нулю. Так, на рис. 48 продемонстрировано, что в зависимости от физиологического и психологического состояния одна и та же собака может показывать результаты, различающиеся практически в 1,5 — 2 раза.
Способность собак разного пола, породы, рабочей специализации различать низкие концентрации запаховых веществ в большей степени определяется не наследственными качествами, а специальным тренингом по выработке стойких сигнальных реакций (посадка, лай, царапание) на специфическое запаховое вещество.
Таким образом, успех применения собак розыскной службы в мероприятиях, где требуется различать запахи разной концентрации, определяется главным образом квалификацией специалистов-кинологов, а не породной или половой принадлежностью животных.
Промеры
Как и у других животных, промеры у собак характеризуются низкой изменчивостью — коэффициент разнообразия колеблется в пределах от 3 до 5%, причем часто животные внутри одной породы
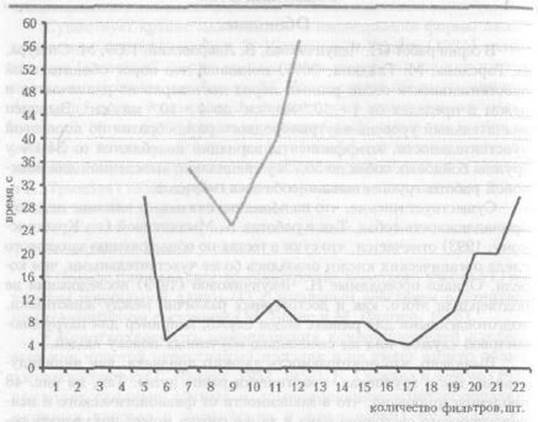
Рис. 48. Продолжительность поиска у одной и той же собаки в разные дни.
различаются больше, чем в среднем между разными породами. Например, разнообразие по высоте в холке сук немецких овчарок в Германии составляет 3,3%, а в России — 5,1%.
В настоящий момент в стандарт большинства пород внесены только два показателя — высота в холке и индекс формата. Поэтому, как правило, наиболее интенсивный отбор проводят по высоте в холке, старясь связать ее значения с уровнем развития рабочих качеств. Показательным примером в этом отношении является история развития в нашей стране породы "немецкая (восточноевропейская) овчарка". Она в полной мере отражает отсутствие четких критериев в оценке конституции и экстерьера. Так, начиная с 1955 года дважды изменялись положения стандартов (в 1964 и 1977 годах), касающиеся значения высоты в холке. В 1964 году границы отбора по высоте в холке были значительно увеличены, поскольку возникло мнение, что
более крупные собаки будут обладать лучшими рабочими качествами в условиях России. Вследствие этого направленного отбора к 1975 году по сравнению с 1955 годом немецкие (восточноевропейские) овчарки достоверно увеличились по высоте в холке (с 68,8 до 70,4 см у кобелей и с 63,8 до 66,4 см у сук). Индекс формата остался неизменным, но возросли грудные промеры, что привело к увеличению индекса массивности. Попытка связать происходившие изменения экстерьера с рабочими качествами показала полное отсутствие связи, т.е. существенные изменения промеров не сопровождались адекватными изменениями рабочих качеств (В. Лавровский, М. Гладких, 1993).
Живая масса
В исследованиях шведских ученых показано, что коэффициент наследуемости живой массы у щенков составляет 0,45 для кобелей и 0,34 для сук. Это означает, что селекция по данному показателю может быть весьма эффективной.
Согласно М. Уиллису (2000), живая масса при одной и той же высоте в холке у разных собак одного пола может колебаться в пределах 37% от средней по породе. Видимо, это характерно для всех пород, которые в стандарте имеют ограничения по высоте в холке. Г. Кайзер (1971) показал, что вес щенка при рождении зависит от живой массы его матери (составляет около 12%, индивидуальные отклонения в пределах 2 — 3%) и средней живой массы по породе. Р. Робинсон в 1973 году предложил следующую формулу для расчета размера помета в зависимости от живой массы суки:
у = 3,32 + 0,36 • х, где х — живая масса суки, кг.
Данное уравнение, по мнению автора, однако, неприменимо к мастифам, сенбернарам и ньюфаундлендам, поскольку они имеют меньшее число щенков в помете, чем ожидаемое при их живой массе. Корреляция между числом щенков в помете и живой массой суки составляет 0,89.
Воспроизводительные качества
В качестве показателей, характеризующих воспроизводительную способность собак, могут быть рассмотрены: интервал между течками,
многоплодие, продолжительность щенности, размер помета, число мертворожденных в помете и т.д.
В исследованиях на 121 суке породы "бигль" показано, что повторяемость интервала между течками составляет 0,43, а наследуемость этого признака — 0,43.
К сожалению, в настоящий момент отсутствуют данные о наследуемости продолжительности щенности у собак разных пород. Тем не менее, в опытах Кржижановского, Студниченко и Малиновского (1975) на 4773 щенных суках разных пород показано, что нет прямой связи между размером суки и продолжительностью ее щенности: для боксеров — 63,5 ± 7,5 дней, для пекинесов — 61,4 ± 7,4 дня.
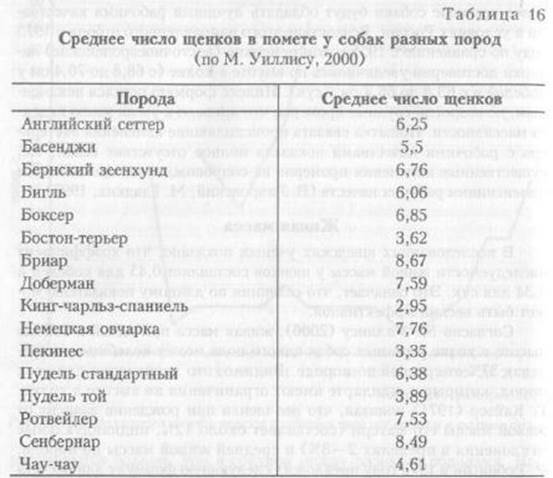
Разные породы собак достоверно различаются по числу щенков в помете. Данные о среднем числе щенков в помете у собак разных пород приведены в табл. 16.
Отметим, что разнообразие собак по числу щенков в помете даже внутри одной породы достаточно велико и находится в пределах от 31,2 до 41,6%, а для басенджи составляет 60,5%, что означает возможное наличие внутри породы групп животных, крайне различающихся по этому показателю, или здесь имеет место отклонение от нормального распределения как следствие специального отбора животных. Размер помета также может изменяться и по годам.
В работах К. Литтла (1949), М. Уиллиса (1976) и Э. Лингсета (1973) показано, что размер помета зависит в большей степени от возраста суки, хотя и может меняться в зависимости от возраста кобеля.
Кроме размера помета большое значение при оценке воспроизводительных качеств собак имеет процент выживания. М. Уиллис (1977) на немецких овчарках выявил следующую закономерность: коэффициент мертворожденных в помете из 1 — 10 щенков составил 3,83%, а в пометах из 11 — 17 щенков — 5,08%. Процент пометов с 10 щенками, где отмечен хотя бы один мертворожденный, остается одним и тем же (12 — 15%), а далее с увеличением числа щенков при рождении пропорционально возрастает. Ф. Гейне и Л. Ван Флек (1976) установили, что новорожденные в помете с весом больше среднего выживают чаще тех, у которых он ниже среднего. Коэффициент корреляции между живой массой новорожденного и смертностью щенков в течение первых шести недель составляет в среднем 0,42. Также у щенка больше шансов на выживание в средних пометах, чем в крупных, — каждый дополнительный щенок сверх среднего размера помета повышает коэффициент смертности в помете на 3,48% в течение первой недели и на 3,43% — в период со второй по шестую неделю. Коэффициент наследуемости живой массы при рождении составляет 10,6%, смертности в первую неделю жизни — 9,5%, смертности в период с 8-го по 42-й день — 8,2%. Ученые Корнельского университета (М. Уиллис, 2000) показали, что влияние кобеля на смертность Щенков в первую неделю жизни на 16,7% выше, чем влияние сук.
Выявление крипторхизма является одной из важных задач при оценке воспроизводительных качеств собак. Т. Паллиг (1953), В. Губер и Е. Шмид (1959) пришли к заключению, что крипторхизм связан с проявлением рецессивного гена, сцепленного с Х-хромосомой. У нормальных самцов и самок Х-хромосомы несут доминантный ген С, обеспечивающий отсутствие аномалий. Поэтому у нормальных самцов их половые хромосомы выражаются как XCY, а у самок — как ХСХС (Е. Меркурьева, 1986). При наличии гена крипторхизма генотип самца будет Xе Y, а самка может быть гетерозиготна по этому гену ХСХС. Отсюда ясно, что распространение крипторхизма в потомстве происходит через самок — носительниц гена С. Т. Пендерграсс и X. Хейес (1975) выявили, что развитие опухолей неопустившихся семенников у крипторхов происходит в 10,9 раза чаще, чем у здоровых кобелей.
Дата добавления: 2016-06-18; просмотров: 2618;
Поиск по сайту
Узнать еще
- ІІ.5.4. Определение фрактальной размерности крестовых клеточных структур
- А). Непосредственные умозаключения: превращение, обращение, противопоставление предикату, по логическому квадрату.
- А. Общего пользования – поступает от разных поставщиков.
- Абзац с выравниванием по центру, шрифт Arial, размер 14 пт, полужирный
- Аллергические реакции могут проявляться в виде слабой тошноты и кишечных спазм, диареи или отеках, размеры которых могут превышать пять сантиметров в диаметре.
- АНАЛИЗ РАЗМЕРНОСТЕЙ И МЕТОД АНАЛОГИЙ
- Атомистическая философия Демокрита - попытка решения проблемы поставленной Парменидом.
- В зависимости от количества и размеров конкрементов желчного пузыря

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
