Классификация рефлексов
Все рефлексы делятся на 2 большие группы: безусловные и условные (приобретенные). Безусловные (врожденные) рефлексы, в реализации которых принимает участие кора головного мозга и подкорковое образование, рассматриваются в главе XV.
Безусловные рефлексы можно классифицировать на группы по ряду признаков.
По месту расположения рецепторов, вызывающих рефлекторный акт: экстерорецептивные (лат. exterus находящийся вне, наружный+ рефлекс) - рефлексы на слуховые, обонятельные, вкусовые, зрительные, механические и термические стимулы. В отличие от рефлексов на раздражение рецепторов внутренней среды экстерорецепторы быстрее формируются (через 5-20 сочетаний), но и быстрее угасают.
Интерорецептивные рефлексы (лат. interior внутренний + capio брать, принимать) - рефлексы, возникающие при раздражении интерорецепторов вследствие изменений условий среды и направленные на сохранение постоянства внутренней среды. Выражаются чаще не в инициации деятельности какого-либо органа, а в изменении характера его функционирования (например, реакции на растяжение полых органов, активация сосудистых рефлексогенных зон); проприрецептивные рефлексы (лат. proprius собственный+ capio брать, принимать) - рефлексы, возникающие при раздражении рецепторов, расположенных в структурах опорно-двигательной системы (суставные, сухожильные, мышечные) и в тканях (механорецепторы, хеморецепторы и др.).
По расположению центрального звена: спинальные (двигательные); бульбарные (глотательные, дыхательные, слюноотделительные); мезэнцефалические (ориентировочные, зрительные, слуховые); диэнцефалические (защитные, пищевые, половые); мозжечковые; корковые.
По локализации эфферентной части: соматические и вегетативные рефлексы.
По характеру ответной реакции: двигательные, секреторные, сосудистые.
По характеру влияния на деятельность эффектора: возбудительные и тормозные.
По биологическому значению: пищевые, оборонительные, половые, локомоторные и др. рефлексы.
Временем рефлекса называют время от момента нанесения раздражения до появления ответной реакции. Оно состоит из времени, которое затрачивается на возникновение возбуждения в рецепторе, времени прохождения по афферентному пути, времени передачи импульсом в ЦНС через последовательный ряд синапсов с афферентного пути на эфферентный, времени передачи возбуждения эфферентному пути и времени возбуждения рабочего органа.
Время проведения возбуждения в ЦНС называется центральным временем рефлекса. Оно тем больше, чем сложнее рефлекторный акт (чем больше промежуточных нейронов участвует в его осуществлении, тем больше происходит синаптических переключений). Установлено, что время рефлекса зависит от силы раздражения: оно тем меньше, чем больше сила раздражения, и, наоборот, время рефлекса тем больше, чем слабее раздражение
Рефлекторная дуга (нервная дуга) — путь, проходимый нервными импульсами при осуществлении рефлекса.
Рефлекторная дуга состоит из:
рецептора — нервное звено, воспринимающее раздражение;
афферентного звена — центростремительное нервное волокно — отростки рецепторных нейронов, осуществляющие передачу импульсов от чувствительных нервных окончаний в центральную нервную систему;
центрального звена — нервный центр (необязательный элемент, например для аксон-рефлекса);
эфферентного звена — осуществляют передачу от нервного центра к эффектору.
эффектора — исполнительный орган, деятельность которого изменяется в результате рефлекса.
Различают:
моносинаптические, двухнейронные рефлекторные дуги;
полисинаптические рефлекторные дуги (включают три и более нейронов).
Полисинаптическая рефлекторная дуга: нервный импульс от рецептора передаётся по чувствительному (афферентному) нейрону в спинной мозг. Клеточное тело чувствительного нейрона расположено в спинальном ганглии вне спинного мозга. Аксон чувствительного нейрона в сером веществе мозга связан посредством синапсов с одним или несколькими вставочными нейронами, которые, в свою очередь, связаны с дендритами моторного (эфферентного) нейрона. Аксон последнего передаёт сигнал от вентрального корешка на эффектор (мышцу или железу).
Понятие введено М. Холлом в 1850 г. В настоящее время понятие рефлекторной дуги не полностью отражает механизм осуществления рефлекса, и в связи с этим Н. А. Бернштейном был предложен новый термин — «Рефлекторное кольцо», в состав которого входит недостающее звено контроля, осуществляемого нервным центром за ходом работы исполнительного органа — т. н. обратной афферентации
Следует отметить, что термин «Рефлекторное кольцо» впервые был использован А. Ф. Самойловым в статье «Кольцевой ритм возбуждения»[1]. Поэтому приоритет в данном словосочетании принадлежит отнюдь не Бернштейну Н. А. и тем более не Анохину П. К., а советскому физиологу, сподвижнику Павлова И. П. и Сеченова И. М., учёному Самойлову А. Ф.
По данным редактора Шестакова М. П., исследователь Бернштейн Н. А. ввёл данный термин в 1935 году, но уже, как было указано, в 1930 году термин «рефлекторное кольцо» использовался в работах Самойлова А. Ф.
Простейшая рефлекторная дуга у человека образована двумя нейронами — сенсорным и двигательным (мотонейрон). Примером простейшего рефлекса может служить коленный рефлекс. В других случаях в рефлекторную дугу включены три(и более)нейрона — сенсорный, вставочный и двигательный. В упрощенном виде таков рефлекс, возникающий при уколе пальца булавкой. Это спинальный рефлекс, его дуга проходит не через головной, а через спинной мозг. Отростки сенсорных нейронов входят в спинной мозг в составе заднего корешка, а отростки двигательных нейронов выходят из спинного мозга в составе переднего. Тела сенсорных нейронов находятся в спинномозговом узле заднего корешка (в дорсальном ганглии), а вставочных и двигательных — в сером веществе спинного мозга.
Простая рефлекторная дуга, описанная выше, позволяет человеку автоматически (непроизвольно) адаптироваться к изменениям окружающей среды, например, отдергивать руку от болевого раздражителя, изменять размеры зрачка в зависимости от условий освещенности. Также она помогает регулировать процессы, протекающие внутри организма. Все это способствует сохранению постоянства внутренней среды, то есть поддержанию гомеостаза.
Во многих случаях сенсорный нейрон передает информацию (обычно через несколько вставочных нейронов) в головной мозг. Головной мозг обрабатывает поступающую сенсорную информацию и накапливает её для последующего использования. Наряду с этим головной мозг может посылать моторные нервные импульсы по нисходящему пути непосредственно к спинальным мотонейронам; спинальные мотонейроны инициируют ответ эффектора.
Рецептивное поле (англ. receptive field) сенсорного нейрона — участок с рецепторами, которые при воздействии на них определённого стимула приводят к изменению возбуждения этого нейрона.
Концепция рецептивных полей может быть применима ко всей нервной системе. Если множество сенсорных рецепторов образуют синапсы c единственным нейроном, они совместно формируют рецептивное поле этого нейрона. Например, рецептивное поле ганглионарной (ганглиозной) клетки сетчатки глаза представлено фоторецепторными клетками (англ.)русск. (палочками или колбочками), а группа ганглионарных клеток в свою очередь создаёт рецептивное поле для одного из нейронов мозга. В итоге к одному нейрону более высокого синаптического уровня сходятся импульсы от многих фоторецепторов; и этот процесс называется конвергенцией.
Понятие о рефлексе впервые возникло в физике Декарта. Декарт развивал общую механическую картину мира и хотел включить в нее также и поведение живых существ. Концепция Декарта была создана в эпоху, когда разные ученые давали материалистические объяснения феноменам, происходящим в природе: например, медик Уильям Гарвей открыл и описал систему кровообращения, в которой эта система выступала как простой механизм с известными к тому времени человечеству элементами - насосом, "трубами" и т.п. Теория Декарта еще более укрепляла принцип материалистического детерминизма, так как Гарвей рассматривал в качестве механизма только внутреннее устройство тела животного, а Декарт перенес этот принцип также и на взаимодействие организмов с внешним миром, т.е., по сути, на психическую деятельность. Декарт считал, что взаимодействие организмов с окружающими телами опосредовано нервной машиной, в которой мозг выполняет роль центра, а от него расходятся "нервные трубки". Согласно его схеме, внешние факторы действуют на концы расположенных в теле нервных "нитей", которые, натягиваясь, открывают клапаны отверстий, ведущих из мозга в нервы. По этим открытым каналам "животные духи" (выражение Декарта) устремляются в соответствующие мышцы, которые в результате "надуваются". Таким образом Декарт утвреждал, что причина двигательного акта не является продуктом мозга или души, а лежит извне, снаружи организма. Работа Декарта была результатом вдохновляющего влияния Гарвея: Декарт писал, что по движениям органов кровообращения "как по первому и самому общему, что наблюдают в животных, можно легко судить и обо всем остальном". Хотя термин "рефлекс" у Декарта отсутствует, ему удалось наметить контуры этого понятия вполне отчетливо: И. П. Павлов считал именно Декарта основоположником понятия "рефлекс" в физиологии и писал, что "Считая деятельность животных, в противоположность человеческой, машинообразной, Декарт... установил понятие рефлекса как основного акта нервной системы".
Предположение о полностью рефлекторном характере деятельности высших отделов головного мозга впервые было развито ученым-физиологом И. М. Сеченовым. До него физиологи и неврологи не решались поставить вопрос о возможности физиологического анализа психических процессов, которые предоставлялось решать психологии.
Далее идеи И. М. Сеченова получили развитие в трудах И. П. Павлова, который открыл пути объективного экспериментального исследования функций коры, разработал метод выработки условных рефлексов и создал учение о высшей нервной деятельности. Павлов в своих трудах ввел деление рефлексов на безусловные, которые осуществляются врожденными, наследственно закрепленными нервными путями, и условные, которые, согласно взглядам Павлова, осуществляются посредством нервных связей, формирующихся в процессе индивидуальной жизни человека или животного.
Учение о рефлексах дало очень многое для понимания самой сущности нервной деятельности. Однако сам рефлекторный принцип не мог объяснить многие формы целенаправленного поведения. В настоящее время понятие о рефлекторных механизмах дополнено представлением о роли потребностей в организации поведения, стало общепринятым представление о том, что поведение животных организмов, в том числе и человека, носит активный характер и определяется не столько возникающими раздражениями, сколько планами и намерениями, возникающими под влиянием определённых потребностей. Эти новые представления получили своё выражение в физиологических концепциях «функциональной системы» П. К. Анохина или «физиологической активности» Н. А. Бернштейна. Сущность этих концепций сводится к тому, что мозг может не только адекватно отвечать на внешние раздражения, но и предвидеть будущее, активно строить планы своего поведения и реализовать их в действии. Представления об «акцепторе действия», или «модели потребного будущего», позволяют говорить об «опережении действительности».
Биологическая концепция рефлекса была сформирована чешским анатомом и физиологом Йиржи Прохазкой (1749-1820). Свои представления о рефлексе Й. Прохазка выразил следующим образом: внешние впечатления, возникающие в чувствительных нервах, быстро распространяются по всей их длине до самого начала. Там они отражаются по определенному закону, переходят на соответствующие им двигательные нервы и по ним очень быстро направляются к мышцам, которые затем производят точные и строго ограниченные движения. Впервые термин «рефлекс» был введен в научный язык Й. Прохазкой. В отличие от Р. Декарта, Й. Прохазка утверждает монистическое представление о нервной системе, в целом относящейся к композиции «общего чувствилища», телесная часть которого локализуется в спинном мозге, а душевная – в головном.
22. Понятие рефлекторной дуги претерпело существенную эволюцию. Классическая рефлекторная дуга имеет три звена: афферентное, центральное и эфферентное.Представление о кольцевой организации рефлекса за счет сигнализации от мышц были завершены в трудах Н.А.Бернштейна (1966) и Ф.В.Бассина (1968), которые дополнили классическую рефлекторную дугу четвертым звеном –афферентацией от эффекторов, в частности, мышечного действия. Вследствие этого рефлекторная дуга трансформировалась в «рефлекторное кольцо».Т.о. структурная основа рефлексарассматривается в настоящее время как кольцевая организация, в которой эффекторы постоянно сигнализируют в соответствующие центры об изменении своего состояния под действием тех или иных раздражителей. И все-таки на основе рефлекторного принципа оказалось трудным объяснить механизмы активной целенаправленной деятельности животных в естественной среде обитания, поиск ими специальных раздражителей внешнего мира.
Все это стимулировало исследователей к новым поискам, которые привели к формированию новых системных представлений об организации физиологических функций и поведения. Исходным пунктом создания теории ФС явились работы П.К.Анохина и сотрудников, проводившиеся в начале 30-х годов по выяснению соотношения центрально-периферических факторов, лежащих в основе компенсаторных приспособлений организма. На основе своих работ, П.К.Анохин высказывает мысль о наличии в механизмах поведения животных аппарата «предсказания» будущего результата действия, а значит о наличии в структуре целостного поведенческого акта особого звена, информирующего ЦНС об успешности достигнутого результата. Открытие в механизмах поведения акцептора результата действия и обратной афферентации позволило рассмотреть поведенческий акт уже не как трехчленную рефлекторную дугу, а как замкнутое циклическое образование с непрерывной обратной информацией об успешности приспособительного действия.
Уже в 1935 году П.К.Анохин сформулировал первое определение ФС: «под ФС мы понимаем круг определенных физиологических проявлений, связанных с выполнением какой-то определенной функции (акт дыхания, акт глотания, локомоторный). Каждая ФС представляет собой до некоторой степени замкнутую систему, протекает благодаря постоянной связи с периферическими органами и в осоСформулировав понятие о ФС, П.К.Анохин тем не менее никогда не отрицал рефлекторный принцип деятельности, но между ними нельзя ставить и знака равенства. ФС имеет специфические и только свойственные системе узловые механизмы для каждого этапа развития действия, свои собственные специфические принципы организации. Т.о. теория ФС выросла из рефлекторной теории в павловском ее понимании, является ее продолжением, но вместе с тем, в известном смысле, и ее отрицанием. Однако отрицание это является диалектическим.
В отличие от рефлекторного принципа, объясняющего «деятельность целого мозга» в процессе приспособительного поведения, теория ФС направлена на изучение целостного организма в его неразрывной связи с окружающей средой
Любая ФС, по мнению П.К.Анохина включает следующие общие, универсальные для различных систем узловые механизмы:
1.полезный приспособительный результат, как ведущее звено ФС,
2.рецепторы результатов,
3.обратную афферентацию, идущую от рецепторов результата в центральные образования ФС,
4.центральную архитектуру, представляющую избирательное объединение ФС нервных элементов различных уровней,
5.исполнительные соматические, вегетативные и эндокринные компоненты, включающие организованное, целенаправленное поведение.
Объединение всех условных механизмов в ФС направлено на достижение результата ее деятельности, полезного для организма в целом. Любое изменение результата, также как и его оптимальное для организма состояние, воспринимается соответствующими рецепторами. Сигнализация, возникающая в рецепторах, поступает в нервный центр и избирательно вовлекает элементы различного уровня в данную ФС для построения исполнительной деятельности, включая различные органы для восстановления потребного для метаболизма результата. Т.о. структурной основой рефлекса является не рефлекторная дуга, а рефлекторное кольцо, состоящее из звеньев (1-6).Анализ структурной основы рефлекса проводится путем последовательного выключения отдельных звеньев рефлекторного кольца. При выключении любого звена рефлекторного кольца рефлекс исчезает. Следовательно, для осуществления рефлекса необходима целостность всех звеньев его морфологической основы.
23. Синапс —специализированная зона контакта между нейронами или нейронами и другими возбудимыми образованиями, обеспечивающая передачу сигналов с сохранением, изменением или исчезновением ее информационного значения.
Если иннервируемая клетка нервная, то синапс называется межнейронным, кроме межнейронных синапсов по месту расположения выделяют :
1. нейроэффекторные (иннервируемая клетка мышечная или железистая)
2. нейрорецепторные (контакт между нейроном и вторично-чувствующей рецепторной клеткой).
• Межнейронные синапсы делятся на:
1. аксосоматические
2. аксодендритические
3. аксо-аксональные в зависимости от того, с какой частью иннервируемой клетки контактирует аксон –соответственно с телом клетки, дендритом, аксоном.
Есть также дендро-дендритические, дендросоматические и сомато-соматические синапсы, но они встречаются редко и функция их не выяснена.
Нейроэффекторные синапсы в зависимости от органа-эффектора бывают нейромышечные, нейрососудистые, нейросекреторные.
• По действию на иннервируемую клетку синапсы делят на :
1.возбуждающие
2. тормозные.
• По механизму передачи существуют
1. химические
2. электрические
3. смешанные.
Структура и механизм действие синапса
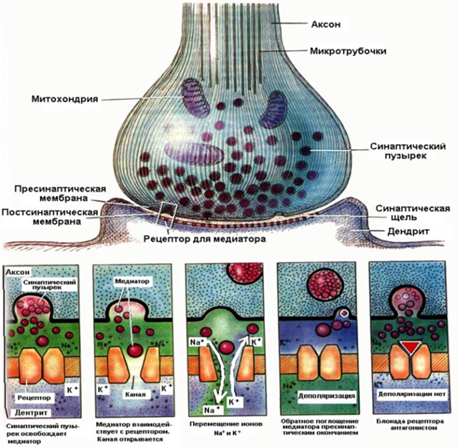
Синаптические процессы в возбуждённом и невозбуждённом синапсе
Контактное взаимодействие обусловлено специфическими контактами мембран нейронов, которые образуют так называемые электрические и химические синапсы.
Электрические синапсы. Морфологически представляют собой слияние, или сближение, участков мембран. В последнем случае синаптическая щель не сплошная, а прерывается мостиками полного контакта. Эти мостики образуют повторяющуюся ячеистую структуру синапса, причем ячейки ограничены участками сближенных мембран, расстояние между которыми в синапсах млекопитающих 0,15—0,20 нм. В участках слияния мембран находятся каналы, через которые клетки могут обмениваться некоторыми продуктами. Кроме описанных ячеистых синапсов, среди электрических синапсов различают другие — в форме сплошной щели; площадь каждого из них достигает 1000 мкм, как, например, между нейронами ресничного ганглия.
Электрические синапсы обладают односторонним проведением возбуждения. Это легко доказать при регистрировании электрического потенциала на синапсе: при раздражении афферентных путей мембрана синапса деполяризуется, а при раздражении эфферентных волокон — гиперполяризуется. Оказалось, что синапсы нейронов с одинаковой функцией обладают двусторонним проведением возбуждения (например, синапсы между двумя чувствительными клетками), а синапсы между разнофункциональными нейронами (сенсорные и моторные) обладают односторонним проведением. Функции электрических синапсов заключаются прежде всего в обеспечении срочных реакций организма. Этим, видимо, объясняется расположение их у животных в структурах, обеспечивающих реакцию бегства, спасения от опасности и т. д. Электрический синапс сравнительно мало утомляем, устойчив к изменениям внешней и внутренней среды. Видимо, эти качества наряду с быстродействием обеспечивают высокую надежность его работы.
Химические синапсы. Структурно представлены пресинаптической частью, синаптической щелью и постсинаптической частью. Пресинаптическая часть химического синапса образуется расширением аксона по его ходу или окончания (рис. 2.19). В пресинаптической части имеются агранулярные и гранулярные пузырьки. Пузырьки (кванты) содержат медиатор. В пресинаптическом расширении находятся митохондрии, обеспечивающие синтез медиатора, гранулы гликогена и др. При многократном раздражении пресинаптического окончания запасы медиатора в синаптических пузырьках истощаются. Считают, что мелкие гранулярные пузырьки содержат норадреналин, крупные — другие катехоламины. Агранулярные пузырьки содержат ацетилхолин. Медиаторами возбуждения могут быть также производные глутаминовой и аспарагиновой кислот. Для синапсов с химическим способом передачи возбуждения характерны синоптическая задержка проведения возбуждения, длящаяся около 0,5 мс, и развитие постсинаптического потенциала (ПСП) в ответ на пресинаптический импульс. Этот потенциал при возбуждении проявляется в деполяризации постсинаптической мембраны, а при торможении — в гиперполяризации ее, в результате чего развивается тормозной постсинаптический потенциал (ТПСП). При возбуждении проводимость постсинаптической мембраны увеличивается.
ВПСП возникает в нейронах при действии в синапсах ацетил холина, норадреналина, дофамина, серотонина, глутаминовой кислоты, вещества Р.
ТПСП возникает при действии в синапсах глицина, гамма-аминомасляной кислоты. ТПСП может развиваться и под действием медиаторов, вызывающих ВПСП, но в этих случаях медиатор вызывает пе-реход постсинаптической мембраны в состояние гиперполяризации. Для распространения возбуждения через химический синапс важно, что нервный импульс, идущий по пресинаптической части, полностью гасится в синаптической щели. Однако нервный импульс вызывает физиологические изменения в пресинаптической части мембраны. В результате у ее поверхности скапливаются синаптические пузырьки, изливающие медиатор в синаптическую щель.
23. В возбуждающих синапсах нервной системы медиатором может являться ацетилхолин, норадреналин, дофамин, серотонин, глугаминовая кислота, вещество Р, а также большая группа других веществ, являющихся, если не медиаторами в прямом значении, то во всяком случае модуляторами (меняющими эффективность) синаптической передачи. Возбуждающие медиаторы вызывают появление на постсинаптической мембране возбуждающего постсинаптического потенциала(ВПСП). Его формирование обусловлено тем, что медиатор-рецепторный комплекс активирует Na- каналы мембраны (а также вероятно и Са-каналы) и вызывает за счет поступления натрия внутрь клетки деполяризацию мембраны. Одновременно происходит и уменьшение выхода из клетки ионов К+ Амплитуда одиночного ВПСП однако довольно мала, и для уменьшения заряда мембраны до критического уровня деполяризации необходима одновременная активация нескольких возбуждающих синапсов. ВПСП, образуемые на постсинаптической мембране этих синапсов, способны суммироваться, т.е. усиливать друг друга, приводя к росту амплитуды ВПСП (пространственная суммация). Растет амплитуда ВПСП и при увеличении частоты поступающих к синапсу нервных импульсов (временная суммация), что повышает число выводимых в синаптическую щель квантов медиатора. Процесс спонтанной регенеративной деполяризации возникает в нейроне обычно в месте отхождения от тела клетки аксона, в так называемом аксонном холмике, где аксон еше не покрыт миелином и порог возбуждения наиболее низкий. Таким образом, ВПСП, возникающие в разных участках мембраны нейрона и на его дендритах, распространяются к аксонному холмику, где суммируются, деполяризуя мембрану до критического уровня и приводя к появлению потенциала действия.
25. В тормозных синапсах обычно действуют другие, тормозные, медиаторы. Среди них хорошо изученными являются аминокислота глицин (тормозные синапсы спинного мозга), гамма-аминомасляная кислота (ГАМК) — тормозной медиатор в нейронах головного мозга. Вместе с тем, тормозной синапс может иметь тот же медиатор, что и возбуждающий, но иную природу рецепторов постсинаптической мембраны. Так, для ацетилхолина, биогенных аминов и аминокислот на постсинаптической мембране разных синапсов могут существовать как минимум два типа рецепторов, и, следовательно, разные медиатор-рецепторные комплексы способны вызывать различную реакцию хемочувствительных рецепторуправляемых каналов. Для тормозного эффекта такой реакцией может являться активация калиевых каналов, что вызывает увеличение выхода ионов калия наружу и гиперполяризацию мембраны. Аналогичный эффект во многих тормозных синапсах имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Возникающий при гиперполяризации сдвиг мембранного потенциала получил название тормозного постсинаптического потенциала (ТПСП). На рис.3.5 показаны отличительные черты ВПСП и ТПСП. Увеличение частоты нервных импульсов, приходящих к тормозному синапсу, также как и в возбуждающих синапсах, вызывает нарастание числа квантов тормозного медиатора, выделяющихся в синаптическую щель, что, соответственно, повышает амплитуду гиперполяризационного ТПСП. Вместе с тем, ТПСП не способен распространяться по мембране и существует только локально. В результате ТПСП уровень мембранного потенциала удаляется от критического уровня деполяризации и возбуждение становится либо вообще невозможным, либо для возбуждения требуется суммация значительно больших по амплитуде ВПСП, т.е. наличие значительно больших возбуждающих токов. При одновременной активации воз-буждаюших и тормозных синапсов резко падает амплитуда ВПСП, так как деполяризующий поток ионов Na+ компенсируется одновременным выходом ионов К+ в одних видах тормозных синапсов или входом ионов СГ в других, что называют шунтированием ВПСП. Под влиянием некоторых ядов может происходить блокада тормозных синапсов в нервной системе, что вызывает безудержное возбуждение многочисленных рефлекторных аппаратов и проявляется в виде судорог. Так действует стрихнин, конкурентно связывающий рецепторы постсинаптической мембраны и не позволяющий им взаимодействовать с тормозным медиатором. Столбнячный токсин, нарушающий процесс освобождения тормозного медиатора, также угнетает тормозные синапсы.
26. Нервный центр-это совокупность нейронов, принимающих участие в осуществлении конкретного рефлекса (мигания, глотания, кашля и т. д.) .
В целом организме при формировании сложных адаптивных процессов происходит функциональное объединение нейронов, расположенных на различных уровнях ЦНС. Такое объединение (нервный центр в широком смысле слова) позволяет осуществлять наиболее адекватное для конкретных условий осуществления рефлекторной деятельности.
Нервные центры обладают рядом характерных функциональных свойств, обусловленных объединением нейронов в нейронные сети и наличием межнейрональных синапс
Основные из этих свойств:
1. Возбуждение в нервных центрах распространяется односторонне-от рецептора к эффектору, что обусловливается свойством химических синапсов односторонне проводить возбуждение от пресинаптической мембраны к постсинаптической.
2. Возбуждение в нервных центрах проводится медленнее, чем по нервному волокну. Это обусловлено замедленным проведением возбуждения через синапс (синаптическая задержка) .
3. В нервных центрах осуществляется суммация возбуждений.
Различают два вида суммации:
•временная или последовательная, если импульсы возбуждения приходят к нейрону по одному и тему же пути через один синапс с интервалом меньше, чем время полной реполяризации постсинаптической мембраны. В этих условиях ВПСП на постсинаптической мембране суммируются и доводят ее деполяризацию до уровня, достаточного для генерации нейроном потенциала действия;
•пространственная или одновременная-наблюдается в том случае, когда импульсы возбуждения поступают к нейрону одновременно через разные синапсы
Трансформация ритма возбуждения -изменение количества импульсов возбуждения, выходящих из нервного центра, по сравнению с числом импульсов, приходящих к нему. Различают два вида трансформации:
•понижающая трансформация, в основе которой, в основном, лежит явление суммации возбуждений (пространственной и временной), когда в ответ на несколько возбуждений, пришедших к нервной клетке, в последней возникает только одно возбуждение;
•повышающая трансформация, в ее основе лежат механизмы умножения (мультипликации), способные резко увеличить количество импульсов возбуждения
Рефлекторное последействие
•заключается в том, что рефлекторная реакция заканчивается позже прекращения действия раздражителя.
Это явление обусловлено двумя причинами:
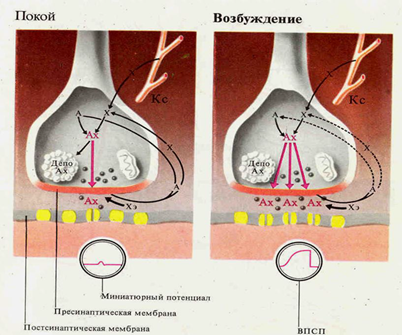
•длительной следовой деполяризацией мембраны нейрона, на фоне которой могут возникать несколько потенциалов действия, обеспечивающих кратковременное рефлекторное последействие;
•пролонгированием выхода возбуждения к эффекторув результате циркуляции (реверберации) возбуждения в нейронной сети типа "нейронной ловушки" (рис. ).
•Возбуждение, попадая в такую сеть, может длительное время циркулировать в ней, обеспечивая длительное рефлекторное последействие. Возбуждение в такой цепочке может циркулировать до тех пор, пока какое-либо внешнее воздействие затормозит этот процесс или в ней наступит утомление. Нервные центры, как и синапсы, обладают высокой чувствительностью к недостатку кислорода.
Нервные центры, как и синапсы, обладают высокой чувствительностью к действию различных химических веществ, особенно ядов. На одном нейроне могут располагаться синапсы, обладающие различной чувствительностью к различным химическим веществам.Поэтому можно подобрать такие химические вещества, которые избирательно будут блокировать одни синапсы, оставляя другие в рабочем состоянии. Это делает возможным корректировать состояния и реакции как здорового, так и больного организма. Нервные центры, как и синапсы, обладают быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми. Нервные центры, как и синапсы, обладают низкой лабильностью. В нервных центрах легко возникает процесс торможения. Нервные центры обладают тонусом, который выражается в том, что даже при отсутствии специальных раздражении, они постоянно посылают импульсы к рабочим органам. Нервные центры, как и синапсы, обладают низкой аккомодационной способностью, т. е. они способны реагировать на раздражающие факторы, медленно нарастающие по силе. Нервные центры обладают пластичностью -способностью изменять собственное функциональное назначение и расширять свои функциональные возможности. Посттетаническая потенциация -явление усиления рефлекторного ответа после длительного ритмического раздражения нервного центра. Оно обусловлено сохранением определенного уровня ВПСП на нейронах центра, что облегчает проведение последующих возбуждений через синапсы.
27. Под торможением в ЦНС следует понимать активный нервный процесс, в результате которого происходит ослабление или полное и длительное выключение возбуждения.
Торможение в ЦНС выполняет две основные функции
•Во-первых, оно координирует функции, т.е. оно направляет возбуждение по определенным путям к определенным нервным центрам, при этом включая те пути и нейроны, активность которых в данный момент не нужна для получения конкретного приспособительного результата.
•(Пример важности этой функции процесса торможения для функционирования организма можно наблюдать в эксперименте с введением животному стрихнина). Стрихнин блокирует тормозные синапсы в ЦНС (в основном, глицинэргические) и, тем самым, устраняет основу для формирования процесса торможения. В этих условиях раздражение животного вызывает некоординированную реакцию, в основе которой лежит диффузная (генерализованная) иррадиация возбуждения.
•Во-вторых,торможение выполняет охранительную или защитную функцию, предохраняя нервные клетки от перевозбуждения и истощения при действии сверхсильных и длительных раздражителей. В ходе эволюции одновременно с процессом возбуждения формировались ограничивающие и прерывающие его механизмы торможения. Классификация центрального торможения по различным признакам:
•По электрическому состоянию мембраны –деполяризационное и гиперполяризационное;
•По отношению к синапсу –пресинаптическое и постсинаптическое;
•По нейрональной организации –поступательное, латеральное (боковое), возвратное и реципрокное.
Торможение в ЦНС (И.М.Сеченов), его виды и механизмы.
Торможение характеризуется снижением или полным прекращением определенной активной деятельности. Вместе с тем, в период торможения активизируются процессы восстановления как в ЦНС, так и на периферии.
Явление центрального торможения было открыто И.М.Сеченовым в 1862 .
Опыт заключался в следующем. У лягушки перерезали ГМ на уровне зрительных бугров и удаляли БП. Измеряли время рефлекса отдергивания задних лапок при погружении их в слабый раствор серной кислоты. Это время является показателем возбудимости нервных центров, поскольку рефлекс осуществляется спинномозговыми центрами. И если на разрез зрительных бугров наложить кристаллик NaCl, то время рефлекса резко удлинится. И.М.Сеченов пришел к заключению, что в таламической области мозга лягушки существует НЦ, оказывающие тормозящие влияния на СМ рефлексы.
Торможение в ЦНС-этоактивныйпроцессвнешнегоподавленияилиослаблениявозбуждения,полногоилидлительноговыключенияопределеннойактивнойдеятельности.
Виды:
• Пресинаптическое
• Постсинаптическое
• Возвратное
• Пессимальное
По нейрональной организациивыделяют
•поступательное,
•латеральное (боковое),
•возвратное,
•реципрокное.
• Поступательное торможение обусловлено включением тормозных нейронов на пути следования возбуждения.
• Латеральное (боковое) торможение .Вставочные клетки формируют тормозные синапсы на соседних нейронах, блокируя боковые пути распространения возбуждения. В таких случаях возбуждение направляется только по строго определенному пути.Именно латеральное торможение обеспечивает, в основном, системную (направленную) иррадиацию возбуждения в ЦНС.
• Возвратное торможение осуществляется вставочными тормозными нейронами (клетками Реншоу). Импульсы от мотонейронов, через отходящие от его аксона коллатерали, активируют клетку Реншоу, которая в свою очередь вызывает торможение разрядов данного мотонейрона. Это торможение реализуется за счет тормозных синапсов, образованных клеткой Реншоу на теле активирующего ее мотонейрона. Таким образом, из двух нейронов формируется контур с отрицательной обратной связью, которая дает возможность стабилизировать частоту разряда мотонейрона и подавлять избыточную его активность. Возвратное торможение имеет значение в деятельности всех отделов ЦНС. Аксоны тормозных нейронов образуют на телах и дендритах возбуждающих клеток нервные окончания, в которых вырабатывается тормозный медиатор (ГАМК в коре БП), в спинном мозге –глицин. Т.е. если тормозные нейроны действуют на те же нервные клетки, которые их активируют, то такая форма торможения получила название возвратного. В этом случае развивающееся торможение бывает тем глубже, чем сильнее было предшествующее возбуждение. Аксоны мотонейронов перед выходом из СМ часто дают один или несколько коллатералей. Они оканчиваются на мотонейронах данного сегмента
Дата добавления: 2016-06-15; просмотров: 12537;
Поиск по сайту
Узнать еще
- Arthropoda..Систематика.Насекомые.Морфология.Классификация.Медицинское значение.
- I. КЛАССИФИКАЦИЯ ПО ИСПОЛЬЗОВАНИЮ.
- I. Классификация углеводов.
- I.4. Классификация групп крови
- III.3. Классификация пород-коллекторов нефти и газа
- IV. Классификация в зависимости от локализации источника кровотечения
- IV. Классификация обязательств РК по международным договорам
- IV.3. Классификация залежей нефти и газа

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
