Математическое моделирование морских экосистем: географо-экологические аспекты
Благодаря усилиям классиков современного естествознания, в ходе истории его развития формировалась качественная модель окружающего мира. Так, В.И. Вернадский заложил основы учения о живом веществе и морской геохимии, А.П. Виноградов начал изучать химический состав микроорганизмов, Н.М. Книпович был пионером рыбопромысловых исследований морей и солоноватых вод, С.В. Бруевич разработал аналитические методы морских гидрохимических работ, сформулировал основы гидрохимии, биогидрохимии и химической динамики морей, Л.А. Зенкевич изучал фауну и биопродуктивность морских вод, А.Б. Скопинцев начал исследования биогенных и органических веществ в водоемах и водотоках, Г.Г. Винберг обращался к вопросам формирования биологической продуктивности морей.
Эти работы послужили методологическим и теоретическим фундаментом начавшихся во всем мире со второй половины ХХ в. регулярных исследований экологического состояния морских экосистем, гидрохимических особенностей формирования сырьевой базы и биопродуктивности природных вод; закономерностей развития химико-биологических процессов трансформации и распада органического вещества (ОВ); механизмов регенерации биогенных субстратов в связи с изучением условий оборачиваемости и круговорота веществ в биосфере, а также способов систематизации и анализа полученной информации.
5.2 Типы математических моделей морских экосистем
Ко второй половине ХХ в. морская экология «созрела» как наука до такой степени, что математическое моделирование состояния морских экосистем стало самостоятельным научным направлением в естествознании. В его рамках Мировой океан рассматривается как сложная динамическая система физических, химических, биологических, геологических и других процессов. Развитие средств вычислительной техники и аппарата прикладной математики привело к интенсивной разработке математических моделей морских экосистем, которые позволили систематизировать полученные знания в различных областях морской науки с целью прогноза и управления состоянием морских водоемов. В настоящее время математические модели морских экосистем, наряду с полевыми наблюдениями в море, стали фундаментом научного понимания природы океана. Для наглядного представления структуры огромного парка типов математических моделей, разработанных в разных странах мира с целью формализации недоступных для прямого инструментального измерения природных процессов или показателей морской среды, В.М. Пищальник составил сводную аналитическую таблицу их видов (рисунок 5.1).
В зависимости от целей моделирования их можно подразделить на имитационные, приуроченные к конкретным бассейнам или районам и разрабатываемые для конкретных целей, и качественные, теоретические, используемые для выяснения общих закономерностей развития и анализа процессов. В имитационных моделях стремятся учесть максимум деталей, а в качественных – минимум, но наиболее важных, поэтому для них главная проблема – выбор приоритетных переменных.
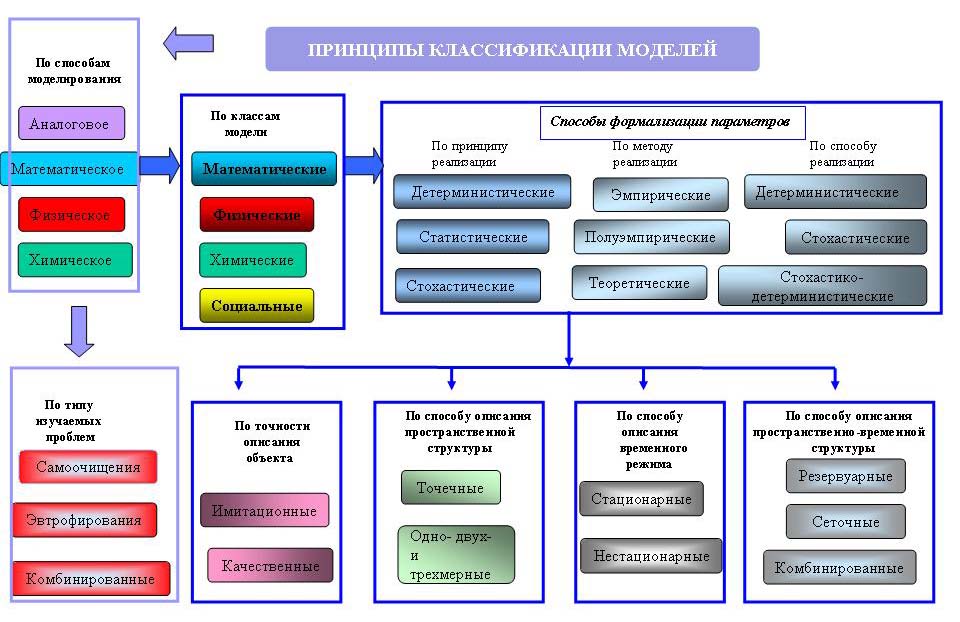
Рисунок 5.1 – Структура парка математических моделей, используемых в эколого-географических исследованиях [17]
По способу реализации модели делятся на детерминистические, в которых для связи между переменными используются функциональные зависимости, и стохастические – на основании статистических связей.
Первые из них используются чаще, так как допускают бесконечное множество компонентов и не учитывают случайных колебаний параметров среды. Они удобны с точки зрения интерпретации результатов. Существуют также стохастико-детерминистические модели, в которых на первом этапе решение ищется детерминистически, а затем, с помощью метода статистических испытаний моделируется изменчивость различных параметров и исследуется реакция решения на эту изменчивость.
По способу представления пространственной структуры явления модели делятся на точечные (параметры сосредоточены в точке), резервуарные (распределение параметров ограничено границами – стенками бокса) и непрерывные (параметры имеют реальное пространственное распределение).
В точечной модели проводится полное интегрирование характеристик по всему объему рассматриваемой области. В резервуарной модели элемент физического пространства, описываемый осредненными по нему (сечение реки, слой океана) характеристиками, называется резервуаром (боксом), а в пространстве состояния (звено пищевой цепи, взвесь, растворенное вещество) – блоком. Передачу свойств от одного резервуара к другому принято называть потоком, а из одного блока в другой – переходом.
В резервуарных моделях рассматриваемый объем разбивается на отдельные резервуары, для каждого из которых учитывается лишь средняя по нему концентрация вещества и строится точечная модель. Это позволяет более или менее грубо учесть пространственную неоднородность и выделить какие-либо структурные объекты. Основным достоинством резервуарных моделей является простота их реализации, хотя в реальной природе трудно выделить репрезентативную систему резервуаров и приписать им надлежащие значения параметров. Кроме того, они чувствительны к малым колебаниям параметров.
В непрерывных моделях осреднения по пространству нет и результатом решения в каждый момент времени служит гладкая кривая (или поле) распределения характеристик. Эти модели обычно сводятся к решению упрощенной системы дифференциальных уравнений термодинамики (двух уравнений движения, уравнения статики, уравнения неразрывности несжимаемой жидкости, уравнения переноса тепла и солей, уравнения состояния), и уравнения для изучаемых характеристик, аналогичных уравнениям баланса тепла и солей.
При реализации задач моделирования химических и биологических характеристик результаты решения системы уравнений термогидродинамики (рассчитанные поля скоростей течения, температуры, солености) подставляют в уравнения переноса рассматриваемых показателей, таким образом представляется возможность исследовать их трансформацию, вызванную химическими, биологическими и биохимическими процессами, совместно с их массопереносом.
Выбор того или иного типа моделей зависит от задачи, стоящей перед исследователем. В этой связи, подчас предпочтение отдается не сложным моделям, а примитивным, из-за их способности дать ответ на конкретно поставленные вопросы.
5.3 Задачи математического моделирования в морской экологии
В комплексе химико-биологических процессов, развивающихся в морской среде, особое внимание при моделировании уделяется механизмам, определяющим трансформацию и биогеохимическую цикличность элементов, входящих в состав живого вещества (С, N, P, S, Si). При этом на первых этапах моделирования внимание акцентировалось на скоростях снижения концентраций химических веществ, утилизации и последовательных превращений субстратов сообществом организмов, а также на скорости и механизме трансформации химических веществ. В результате решались важные практические задачи.
1. Получали количественную информацию об изменениях во времени и пространстве химических и биологических характеристик морской экосистемы в зависимости от интенсивности воздействия на нее факторов среды обитания (температура, освещенность, прозрачность, водный режим, биогенная нагрузка).
2. Оценивалась способность морской экосистемы к самоочищению, и разрабатывались рекомендации для создания наиболее эффективных условий для этого процесса.
3. Составлялся баланс веществ в природных водах с учетом обменных потоков на границах раздела вода – атмосфера, вода – дно, а также поступления веществ с водами притоков и атмосферными осадками и потери веществ при выносе их водными массами из водоема.
4. Устанавливалась роль естественных и антропогенных процессов в круговороте биогенных веществ в морских экосистемах и на этом основании разрабатывались мероприятия по охране водных ресурсов, оценивался запас природных вод, потенциал их качества, определялись способы контроля загрязнения и механизмы очистки вод.
5. Оценивался запас биогенных элементов в природных водах и его пространственная и временная изменчивость под влиянием процессов их потребления планктонными организмами и регенерации при деструкции ОВ. Исследовались основы первичного продуцирования в водоемах и их биологической продуктивности на высших трофических уровнях.
6. Изучались поведение и распределение популяций, сообществ организмов в морской среде, воспроизводились особенности вертикальной неоднородности и горизонтальной мозаичности распределения гидробионтов, устанавливалась роль хищников в регуляции фотосинтетической активности фитопланктона.
7. Изучалась вертикальная структура сообществ микроорганизмов – биомасс разных групп фито- и зоопланктона, детрита, в тропической зоне океана, в пелагиали Японского моря, в пелагиали Перуанского апвеллинга.
8. Исследовались причины активизации (вспышек) развития биологических сообществ, инвазии новых для экосистем видов организмов (нововселенцев), цветения определенных групп водорослей.
В настоящее время актуальными задачами математического моделирования биогеохимических процессов в морских экосистемах являются: изучение скоростей химико-биологических процессов, круговорота веществ в природных водах, условий формирования биологической продуктивности водоемов, оценка балансов соединений органогенных элементов в водной среде, а также комплексное изучение процессов химического обмена на границах раздела вода – атмосфера и вода – дно и химико-биологической трансформации веществ в водной среде и в донных осадках.
При изучении комплекса гидродинамических процессов, развивающихся в морской среде, основное внимание при математическом моделировании состояния морских экосистем уделяется задачам горизонтального и вертикального переноса загрязняющих веществ, установлению условий формирования и распада скоплений морских организмов (фито- и зоопланктона, пелагических и донных промысловых объектов), перераспределения жизни по вертикали.
1.4 Модели биохимических процессов в морских экосистемах
Согласно существующим представлениям о характере процесса обобщения данных экологических исследований на первом его этапе преобладает математическая обработка информации, на втором – моделирование, а на третьем – создание математической теории. Уровень математизации морской экологической науки к 80-м гг. ХХ в. соответствовал первому этапу, хотя первые робкие шаги в рамках второго этапа были сделаны еще в начале ХХ в. – модель кислородного режима рек (Стритера-Фелпса) и взаимоотношений типа хищник-жертва (Волтерра-Лотка), а в мире имелось около 150 математических моделей водоемов и водотоков различной сложности.
Более сотни из этого количества были разработаны для озер и водохранилищ, из них треть создали отечественные авторы. Моделировались экосистемы Великих Американских озер, а также Братского, Иваньковского, Рыбинского, Можайского, Зейского, Днепровских и других водохранилищ. При этом, кроме задач эвтрофирования водоемов, оценок потоков энергии, и продуктивности, решались вопросы влияния периодического затопления суши на экосистему, массового «цветения» водохранилищ. Большинство из этих моделей являются точечными корреляционно-регрессионными и точечными или двухрезервуарными моделями первичного продуцирования и бюджета фосфора как лимитанта и фактора эвтрофирования водных систем. Имитационные модели занимали только 10 % в общем числе выполненных математических разработок и численных экспериментов.
С помощью значительного количества разработанных моделей к 80-му г. исследовались экосистемы континентальных источников, рек, ручьев, а также временных водоемов и прудов, как самых простых объектов для моделирования, и имелись только единичные модели, в которых пытались описать состояние экосистем морских водоемов или отдельных их участков.
К началу ХХI в. в мире уже было разработано достаточно много имитационных моделей, способных аналитически и численно исследовать происходящие в водных экосистемах изменения важнейших химических и биологических характеристик. Их обстоятельный обзор представлен в многочисленных литературных источниках.
Тем не менее, большинство моделей, касающихся вопросов трансформации веществ в морской среде под влиянием биохимических процессов, являются точечными или резервуарными. В пространственных моделях этим вопросам уделяется меньше внимания, и акцент делается больше на анализе пространственно-временной изменчивости гидрохимических и биологических компонентов водных экосистем.
В отличие от моделей распределения гидрофизических показателей состояния морских экосистем, основанных на решении системы стандартных дифференциальных уравнений, в биохимических моделях такой системы уравнений нет. Существуют лишь утвердившиеся способы параметризации моделируемых процессов. При этом чрезвычайно важными оказываются соотношения пространственных и временных масштабов гидрофизических процессов, в рамках которых они способны повлиять на формирование пространственного распределения гидрохимических и биологических показателей. По определению А.С. Монина пространственным неоднородностям присущи типичные времена производящих их процессов (таблица 5.1):
– мелкомасштабным (доли мм – десятки метров) – от 10-3 с. до
десятков часов;
– мезомасштабным (сотни метров – километры) – от часов до суток;
– синоптическим (десятки и первые сотни километров) – от суток до месяцев;
– глобальным (тысячи и десятки тысяч километров) – от лет до сотен лет.
Таблица 5.1 – Процессы, влияющие на распределение и изменчивость химических элементов в океане [Монин, 1982]
| Процесс | Масштаб (сек.) | Время действия | Источник |
| Газообмен океан-атмосфера | 101 | Десятки секунд | Emerson, 1995 |
| Гидролиз газов и процессы в карбонатной системе | 101 | Десятки секунд | Emerson, 1995 |
| Физиологические ритмы морских организмов | 105-6 | Часы-месяц | Рудяков, 1986 |
| Адвективный и турбулентный перенос (поверхностные воды) | 106-7 | Сутки-год | Монин и др., 1974 |
| Химико-биологические | 106-7 | Сутки-год | Монин и др., 1974 |
| Антропогенный СО2 | 109 | 30-40 лет | Gruber et al.1996 |
| Окисление стойкого водного гумуса (В0В) | 1011 | 2000 лет | Скопинцев и др., 1979 |
| Адвективный и турбулентный перенос (глубинные воды) | 1011 | 1000 лет | Монин и др., 1974 |
| Седиментация | > 1012 | > 10000 лет | Монин и др., 1974 |
Очевидно, что на любом масштабе времени в формировании полей гидрохимических показателей активно влияют несколько процессов, что необходимо учитывать при разработке математической модели. При этом в верхнем слое океана, где значение химико-биологических процессов максимально, каждому процессу соответствует определенное характерное время и характерные величины концентраций компонентов. С увеличением глубины такая закономерность нарушается. Содержание живых организмов и ОВ в этом случае снижается экспоненциально, а концентрации биогенных веществ (например, фосфатов) растут. При этом характерные масштабы изменения концентраций растворенных неорганических форм биогенных элементов, а также их взвешенных, органических форм, в том числе в составе живых организмов различаются более, чем на 4 порядка. Таким образом, способ параметризации химико-биологических процессов должен быть связан с масштабами изменчивости концентраций форм химических элементов в выбранном объекте моделирования и пространственно-временным масштабом решаемой задачи.
Под химико-биологическими процессами подразумевается совокупность процессов синтеза ОВ из неорганических соединений, переноса его по трофическим цепям, выделений в виде органических и неорганических форм из живых и мертвых организмов, автолиза взвеси и минерализации ОВ. Процесс синтеза и распада ОВ протекает в рамках классического стехиометрического уравнения Редфилда с учетом обмена веществом между различными организмами в пределах экосистемы.
При моделировании потребления биогенных элементов в процессе синтеза органического вещества (фотосинтезе) принимаются во внимание следующие положения:
– удельная скорость фотосинтеза прямо пропорционально связана с ростом биомассы фитопланктона и скоростью потребления биогенных элементов;
– средний размер клеток фитопланктона и их химический состав одинаковы и постоянны; автотрофный рост прекращается при нулевых значениях освещенности и концентрации биогенных элементов.
При этом учитывается сложная, неоднозначная зависимость интенсивности фотосинтеза от интенсивности и продолжительности освещенности, концентрации биогенных элементов в среде и в клетке, температуры и солености воды, размера и вида растения
Большинство моделей экосистем при описании фотосинтеза опирается на концепцию лимитирующего фактора, управляющего скоростью этого процесса. В 1840 г. Либихом был сформулирован закон, в соответствии с которым скорость фотосинтеза лимитируется тем химическим элементов, отношение концентрации которого в среде к концентрации его в морских организмах минимально. В 1905 г. Блэкман расширил этот закон концепцией лимитирующего фактора, а в 1911 г. Шелфорд сформулировал более общий принцип толерантности, заключающийся в том, что выносливость организма (вида) определяется как максимумом, так и минимумом диапазона изменений экологического фактора. При моделировании морских экосистем в качестве лимитанта обычно используют один из биогенных элементов азот, фосфор, реже кремний. Но в отличие от закона Либиха в этих моделях обычно фигурирует только концентрация лимитанта в воде, но не в организмах.
При моделировании выделения соединений биогенных элементов из автотрофных организмов учитывается поступление этих веществ в морскую среду в процессе дыхания (неорганические соединения), эксудации (потеря органического вещества в процессе жизнедеятельности морских водорослей, которая может достигать 40% от дневной чистой продукции) и при отмирании.
При моделировании трансформации биогенных соединений гетеротрофными организмами учитывается их потребление автотрофами (фитопланктоном) и образование мертвого органического вещества (детрита) зоопланктоном, выделение этих соединений при дыхании, метаболизме и отмирании зоопланктона в виде растворенных неорганических и взвешенных органических форм, а также при выедании одних гетеротрофных организмов другими.
1.5 Гидроэкологическая модель трансформации органогенных элементов
После анализа существующего опыта изучения природы морских водоемов методами математики, для решения задач морской экологической географии была выбрана имитационная боксовая гидроэкологическая модель трансформации органогенных элементов в морской среде, разработанная в конце ХХ в. А.В. Леоновым. Современный вариант модели учитывает характеристики загрязнения (нефтепродукты, пестициды, тяжелые металлы, фенолы), продукцию фитопланктона и концентрации веществ в донных осадках. Отдельные его блоки использовались при исследовании экосистемы Охотского моря, его залива Анива и северо-западного шельфа Черного моря .
В выбранной модели, для изучения режима трансформации НП в морской экосистеме за основу взяты не численность или биомассанефтеокисляющих бактерий,а их биохимическая активность–способность осуществлять цикл процессов, включающий потребление пищевых субстратов, выделение продуктов жизнедеятельности, образование детрита. Модель описывает взаимосвязанные биогеохимические циклы таких элементов, как N и P, а также включает описание скоростей трансформации Si, растворенного органического С и кислорода в двухслойной водной экосистеме. Таким образом, рассчитывается динамика концентраций DOC, О2, N-, Р- и Si-содержащих веществ при их биотрансформации и развитии процессов обмена веществ через границы раздела вода-воздух и вода-дно.
Модель учитывает следующие соединения Р, N и Si: детритный Р, растворенные неорганический и органический Р, органический, аммонийный, нитритный и нитратный N , азот мочевины и свободный азот, неорганический, органический и детритный Si .
Воспроизводимая моделью биотрансформация соединений органогенных элементов осуществляется сообществом микроорганизмов: гетеротрофные бактерии потребляют органические соединения и в процессе метаболизма образуют пул минеральных веществ; фитопланктон утилизирует минеральные вещества и формирует запас органических веществ в водной среде; зоопланктонные организмы регулируют динамику организмов сообщества и своей активностью влияют на развитие продукционно-деструкционных процессов.
Фактически модель воспроизводит переходные процессы и описывает реакцию водной экосистемы на изменения условий среды обитания или на изменение хотя бы одного из учитываемых в модели факторов (водный режим, температура, освещенность водной среды, биогенная нагрузка). Она содержит 226 уравнений, описывающих:
– изменение концентраций исследуемых компонентов;
– удельные скорости потребления органогенных веществ гетеротрофными бактериями, фито- и зоопланктоном;
– функции и коэффициенты для коррекции констант скоростей потребления субстратов гидробионтами на температуру и освещенность;
– удельные скорости метаболических выделений организмов и их выделительные активности;
– удельные скорости отмирания организмов;
– суммарные скорости изменения биомасс микроорганизмов за счет взаимодействий химических и биологических компонентов рассматриваемого сообщества;
– суммарные скорости изменения концентраций биогенных веществ в водной среде и седиментах;
– скорости поступления веществ из внешних источников (с атмосферными осадками и из рассредоточенных источников).
При расчете скоростей изменения концентраций органогенных веществ за счет горизонтального и вертикального переноса учитывается:
· поступление указанных веществ в рассматриваемые акватории моря извне с водами притоков;
· пополнение запаса веществ за счет вертикального обмена с нижележащим слоем;
· перенос веществ из соседних акваторий в пределах морской экосистемы и их потери при выносе водным потоком через внешние границы морской экосистемы.
Блок биохимической трансформации НП нефтеокисляющими бактериями в модели включает 10 уравнений, описывающих:
· изменение концентраций НП и биомассы нефтеокисляющих бактерий;
· суммарное и раздельное потребление субстратов (НП и DOC) нефтеокисляющими бактериям;
· функцию активности нефтеокисляющих бактерий в зависимости от температуры;
· удельные скорости выделения продуктов метаболизма и отмирания биомассы нефтеокисляющих бактерий.
Таким образом, модель описывает внутригодовую динамику химико-биологических показателей состояния водной среды, мгновенные скорости процессов, ответственных за изменение концентраций веществ, внутренние и внешние потоки веществ в разных акваториях исследуемой экосистемы, а также время оборота всех рассматриваемых в модели химических и биологических компонентов.
Вопросы для самоконтроля:
1. Перечислить математические модели, используемые в эколого-географических исследованиях
2. Перечислить задачи математического моделирования в морской экологии
3. Принципы классификации моделей, используемых в экологии
4. Процессы, влияющие на распределение и изменчивость химических элементов в океане
5. Гидроэкологическая модель трансформации органогенных элементов
Литература: [10; 11; 16, 17]
ПРАКТИЧЕСКАЯ РАБОТА № 6
ПРЕДСТАВЛЕНИЕ ОКЕАНОГРАФИЧЕСКИХ ДАННЫХ
Цель занятия:Изучить методику отображения океанографических данных (3 основных способа составления линейных диаграмм).
Задание:
1. Изучить методику отображения океанографических данных (3 основных способа составления линейных диаграмм)
2. Построить графики, диаграммы, карты распределения океанографических параметров по данным в таблицах
3. Выполнить анализ полученных результатов
Материалы и оборудование:Теоретические материалы, таблицы, компьютер.
Дата добавления: 2021-11-16; просмотров: 236;
Поиск по сайту
Узнать еще
- Агроэкологические аспекты взаимодействия ходовой части тракторов и автомобилей с почвой
- АНАТОМО-ФИЗИЗИОЛОГИЧЕСКИЕ АСПЕКТЫ ПОТРЕБНОСТИ ВЫДЕЛЯТЬ
- Анатомо-физиологические аспекты занимающихся различного возраста и квалификации
- Анатомо-физиологические аспекты потребностей человека. Человек как предмет изучения анатомии и физиологии
- Андеррайтинг морских судов
- Ареалы морских животных и растений. Реликты фауны.
- Аспекты в соционике
- Аспекты использования стоимости капитала

Публикации по технике и механике

Публикации по биологии

Публикации по информатике

Публикации по строительству

Публикации по физике

Публикации по химии

Публикации по электронике

Публикации по искусству

Публикации по географии

Публикации по медицине
